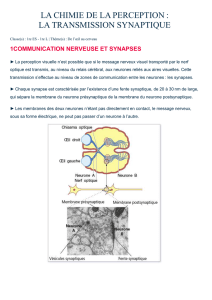
2015-2016 Le système nerveux

51
4.8 Les mécanismes cellulaires de l’apprentissage
D’après un modèle généralement admis, les cellules nerveuses activées en même temps lors de
l’apprentissage forment des réseaux neuronaux spécifiques dans le SNC. Les neurones impliqués dans
le processus d’apprentissage activent des synapses dites silencieuses qui étaient peu utilisées
auparavant, ou forment de nouvelles synapses avec leur cellule voisine. Parallèlement, les synapses
des neurones inutilisés sont mises hors service ou même éliminées.
Figure 47 : Connexions neuronales. A Formation et B activation d’une trace mnésique
En plus de l’élaboration et du démantèlement des connexions cellulaires, l’apprentissage modifie aussi
l’efficacité de la transmission synaptique. Si une synapse n’est pas utilisée pendant une longue
période, une faible dépolarisation de son neurone présynaptique déclenche un PPSE si petit dans le
neurone postsynaptique qu’aucun potentiel d’action n’est généré. Ce n’est que lorsque les potentiels
d’action apparaissent à une fréquence élevée à la suite d’une forte dépolarisation dans le neurone

52
présynaptique qu’un PPSE élevé est généré dans le neurone postsynaptique et que la stimulation est
transmise sous forme de potentiels d’action. Par la suite, une faible stimulation du neurone
présynaptique suffit à déclencher un PPSE assez fort pour former des potentiels d’action. L’efficacité
de la transmission synaptique est renforcée lorsqu’une synapse participe avec succès à l’activation du
neurone postsynaptique. Ce phénomène s’appelle la potentialisation à long terme.
La potentialisation à long terme s’effectue principalement dans les synapses excitatrices qui sécrètent
le neurotransmetteur glutamate. Peu de récepteurs au glutamate sont présents initialement dans la
membrane postsynaptique, rendant ainsi l’efficacité de la transmission synaptique très faible. Lorsque
la sécrétion du neurotransmetteur a suffi pour déclencher un PPSE élevé et, par conséquent, un
potentiel d’action postsynaptique, c’est que la synapse a été impliquée avec succès dans l’activation du
neurone postsynaptique. Une cascade de signalisation est déclenchée dans le neurone activé,
provoquant la formation de récepteurs au glutamate supplémentaires dans la membrane
postsynaptique. En outre, d’autres synapses se forment entre les deux neurones. Ce phénomène
nécessite une activation spécifique de la biosynthèse protéique. Le neurone postsynaptique produit des
médiateurs chimiques qui renforcent la synthèse et la sécrétion des neurotransmetteurs dans le neurone
présynaptique.
Figure 48 : Modifications synaptiques causées par le processus d’apprentissage
A Nombre de récepteurs B Quantité de neurotransmetteurs C Nombre de synapses
Ces divers changements lors de l’apprentissage aboutissent à chaque fois à des réseaux neuronaux
spécifiques dans le système nerveux central. Les neurones concernés sont connectés entre eux par des
synapses excitatrices.

53
Des potentialisations à long terme peuvent être aussi déclenchées lorsque des synapses excitatrices
sont actives en même temps sur un neurone et parviennent ensemble é l’activer. C’est le cas, par
exemple, dans les neurones de l’hippocampe, qui jouent un rôle important lors de l’apprentissage et de
la formation de la mémoire. La potentialisation à long terme des synapses qui transmettent
simultanément des neurotransmetteurs à un neurone permet d’interconnecter tous les neurones
impliqués dans un réseau neuronal.
Ce modèle permet d’expliquer la formation des associations. Si un groupe de synapses transmet
l’information de l’image d’un merle et un autre groupe transmet les informations du chant du merle à
un neurone – dans l’hippocampe par exemple -, l’activation simultanée de ces synapses augmente
l’efficacité de la transmission synaptique et les synapses se connectent entre elles, formant ainsi un
réseau neuronal avec les neurones correspondants. A partir de ce moment, non seulement l’image,
mais aussi le chant – même s’il est perçu seul – aboutissent à la perception du merle. L’apprentissage
associatif (figure 49) a permis de combiner les deux stimuli sensoriels. Les informations de l’image
d’une mésange et le chant d’un merle ne conduisent pas à une telle association, car elles n’arrivent en
même temps qu’occasionnellement.
Figure 49 : L’apprentissage associatif
Plusieurs processus d’apprentissage consistent à élargir les réseaux neuronaux dans le cerveau par la
formation d’associations. Ce qui vient d’être appris est d’autant plus facilement ancré dans le cerveau
que le nombre d’associations avec ce qui est déjà connu est important, et que le nombre de synapses
activées durant l’apprentissage est grand. Cela signifie qu’on apprend plus facilement lorsqu’on sait
déjà quelque chose. L’apprentissage est donc un processus d’autoconsolidation.
L’exercice, c’est-à-dire l’utilisation des synapses et des neurones récemment connectés, renforce
l’efficacité de la transmission de l’excitation dans ces circuits nerveux. Par conséquent, l’accès aux
connaissances est facilité et accéléré. A l’inverse, le renforcement d’une synapse est annulé lorsque les

54
circuits nerveux correspondants ne sont pas utilisés pendant une longue période et qu’on n’a pas utilisé
ce qui a été appris. Ces synapses peuvent disparaître et devront être reformées.
4.9 La mémoire
Au sens où nous l’entendrons ici, la mémoire est la capacité de stocker et de rappeler les informations
enregistrées, les souvenirs. Ces informations sont le résultat de processus d’apprentissage conscients
ou inconscients.
Toutes les informations reçues continuellement par les organes sensoriels, transformées en impulsions
nerveuses et transmises au cerveau par les neurones, ne sont pas stockées dans la mémoire. La plupart
des informations ne sont conservées que brièvement. Par exemple, un numéro de téléphone cité n’est
gardé en mémoire que le temps de le composer. Par contre, d’autres informations, comme sa propre
date de naissance, sont disponibles durant des dizaines d’années. Le modèle modal de la mémoire
distingue différents types de mémoires.
Figure 50 : Modèle modal de la mémoire
La mémoire sensorielle permet d’obtenir une très grande quantité d’informations par les organes
sensoriels, mais ne peut les stocker que pendant quelques fractions de seconde. La plupart des
informations sont très rapidement perdues. Seules celles qui attirent notre attention sont transférées
dans la mémoire de travail. Là, ces informations sont retenues pendant quelques secondes ou quelques
minutes. La répétition de l’information renforce la durée de stockage.
La capacité de la mémoire de travail est cependant très faible. C’est pourquoi le cerveau exécute un
important travail de traitement pour centraliser les informations dans des unités plus grandes. Ainsi, la
quantité globale d’information à stocker est compactée et davantage d’informations peuvent être
emmagasinées. Par exemple, le stockage des quatre lettres V, E, N et T nécessite quatre places de

55
stockage. Par contre, si les lettres sont réunies pour former le mot « VENT », une seule place est
nécessaire.
Quelques informations sont gardées en mémoire durant toute la vie : les mots de la langue maternelle,
les souvenirs d’enfance. Ces informations sont conservées dans la mémoire à long terme. Des
répétitions multiples, ainsi que de l’exercice, sont nécessaires pour transférer les nouvelles
informations dans la mémoire à long terme et les conserver. Le degré d’ancrage dans la mémoire
augmente avec la signification des informations et la possibilité de les relier aux autres informations
déjà sauvegardées.
Le type de stockage est modifié lors du transfert à la mémoire à long terme. Alors que les informations
de la mémoire de travail consistent en influx nerveux, ou activité cérébral, les informations de la
mémoire à long terme sont stockées sous forme de connexions nerveuses, ou de structures
cérébrales. Le transfert entre ces formes de stockage nécessite du temps et du repos. Il a lieu de
préférence lors du sommeil. Le stress rend plus difficile le stockage des informations dans la mémoire
à long terme.
Il existe différents types de mémoire à long terme en fonction des différents contenus : la mémoire
déclarative, aussi appelée la mémoire explicite, contient des informations qui peuvent être rappelées
consciemment et être exprimées avec des mots. On la divise en mémoire épisodique, qui stocke des
souvenirs individuels, et en mémoire sémantique, qui contient des connaissances théoriques ou
factuelles.
La mémoire non déclarative, ou mémoire implicite, est largement utilisée de manière inconsciente.
On distingue la mémoire procédurale, qui concerne principalement des séquences de gestes, par
exemple faire du vélo, et la mémoire émotionnelle, qui peut exercer une influence inconsciente
considérable sur le comportement de l’individu concerné, comme la peur « apprise » au cours
d’expériences désagréables.
Depuis longtemps, on s’est aperçu qu’il n’y avait par une aire bien délimitée du cerveau responsable
de la mémoire. Les analyses des personnes ayant des lésions cérébrales en ont fourni les premières
preuves. Elles ont montré qu’une perte de mémoire était toujours partielle et dépendait de la région
cérébrale atteinte.
Les hippocampes sont des structures essentielles pour le stockage des connaissances. Un patient qui
souffrait de crises d’épilepsie sévères a été traité par l’ablation d’une partie du cerveau, dont les deux
hippocampes. Après l’opération, plus aucune crise d’épilepsie n’apparut chez ce patient et il ne
présentait aucun déficit dans les tests intellectuels. Cependant, il n’était plus en mesure de stocker des
informations déclaratives. Il ne pouvait par exemple retenir un nombre à trois chiffres quand le récitant
constamment. S’il était dérangé, il oubliait non seulement le nombre, mais aussi l’exercice. En
revanche, il était resté capable d’améliorer sa dextérité manuelle par l’exercice. Sa mémoire
procédurale n’était pas atteinte.
Les hippocampes sont reliés avec les diverses aires associatives sensorielles dans les deux directions
par des circuits nerveux, indiquant leur importance pour la mémoire déclarative. On assume
aujourd’hui que les informations déclaratives sont stockées dans les hippocampes durant la journée à
l’état éveillé et qu’ensuite, durant le sommeil, elles sont stockées définitivement dans les différentes
aires associatives.
Le cervelet et les ganglions de la base jouent un rôle important pour la mémoire procédurale, et les
amygdales pour la mémoire émotionnelle.
 6
7
8
6
7
8
1
/
8
100%