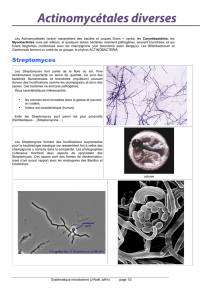
Les actinomycètes

RÉPUBLIQUE ALGÉRIENNE DÉMOCRATIQUE ET POPULAIRE
MINISTÈRE DE L’ENSEIGNEMENT SUPÉRIEUR ET DE LA RECHERCHE SCIENTIFIQUE
N° d'ordre : Année 2014
Université Aboubakr Belkaïd-Tlemcen
Faculté des Sciences de la Nature et de la Vie, des Sciences de la Terre et de l’Univers
Département de Biologie
Laboratoire des Produits Naturels (LAPRONA)
Laboratoire de l’Unité Molécules de Communication et Adaptation des Micro-organismes
MNHN (Paris)
THÈSE
Présentée
par
Mr. BELYAGOUBI Larbi
Pour l’obtention d’un Doctorat en Biologie
Option : Substances Naturelles, Activités Biologiques et Synthèse
*********************
Antibiotiques produits par des bactéries
(actinomycètes et bactéries lactiques) issus de
différents écosystèmes naturels Algériens.
*********************
Soutenue le : Jeudi 26 Juin 2014
Devant le jury composé de :
Année universitaire : 2013-2014
Président : M
r
Chabane Sari D. Professeur Université de Tlemcen
Examinateurs : M
r
Kihal M. Professeur Université d’Oran
M
me
Zirah S.
Maître de conférences
MNHN, Paris
M
me
Bekhechi C.
Maître de conférences
Université de Tlemcen
Directeur de thèse : M
r
Abdelouahid D. E.
Professeur Université de Tlemcen
Co-directeur de thèse :
M
me
Rebuffat S. Professeur MNHN, Paris

i
Remerciements
Avant toute chose, je tiens à remercier «Allah» qui m’a donné la force et la volonté
pour terminer ce modeste travail.
Les travaux présentés dans cette thèse ont été effectués au Laboratoire des Produits
Naturels (LAPRONA) du département de Biologie, de la Faculté des Sciences de la
Nature et de la Vie, des Sciences de la Terre et de l’Univers (SNV/STU), de l’Université
Abou Bakr Belkaïd de Tlemcen. La grande partie de cette étude a été réalisée au sein de
l’unité Molécules de Communication et Adaptation des Micro-organismes, UMR 7245
CNRS/MNHN (anciennement Chimie et Biochimie des Substances naturelles, UMR
5154) dirigée par Madame le professeur Sylvie REBUFFAT. Je tiens à lui exprimer mes
sincères remerciements pour m’avoir permis d’effectuer ces travaux dans cette unité.
L’encadrement scientifique de ce travail a été assuré par Monsieur Djamel Eddine
ABDELOUAHID, Professeur de microbiologie à la Faculté des SNV/STU de Tlemcen.
Je tiens vivement à lui exprimer ma profonde reconnaissance et gratitude pour sa
disponibilité, sa patience, sa compréhension, ses qualités humaines et ses intérêts portés
pour mon sujet de recherche. Je le remercie de m’avoir fait confiance et d’avoir été présent
aussi souvent que possible malgré ses tâches pédagogiques. Son soutien permanent et son
dynamisme m’ont permis d’avancer plus loin dans mes recherches.
Je remercie très chaleureusement ma co-directrice de thèse, Madame Sylvie
REBUFFAT, professeur au MNHN, de l’unité Molécules de Communication et
Adaptation des Micro-organismes, pour avoir accepté de m’accueillir au sein de son
laboratoire pour réaliser des expériences dans ma thèse dans d’excellentes conditions, pour
son soutien et nos conversations constructives.
Je souhaite remercier particulièrement, Madame Séverine ZIRAH, maître de
conférences au MNHN, pour sa grande contribution à ces travaux de recherche, pour son
aide précieuse, pour ses compétences scientifiques, pour sa confiance et ses conseils
scientifiques précieux, pour son soutien et sa gentillesse, pour nos nombreuses discussions
scientifiques, pour ses encouragements et sa bonne humeur.
Je tiens à remercier Monsieur le Professeur Daoudi CHABANE SARI, Directeur
du laboratoire des « Produits Naturels », pour m’avoir fait l’honneur d’accepter la
présidence du jury. Qu’il trouve ici mes sincères impressions de gratitude et de respect.
Mes remerciements vont aussi à Monsieur Mebrouk KIHAL, Professeur à
l’Université d’Oran, d’avoir ménagé son temps pour juger et critiquer ce travail. Je suis
particulièrement reconnaissant et honoré par sa participation au jury de cette thèse.

ii
Que Madame Séverine ZIRAH, maître de conférences au MNHN, trouve ici
l’expression de ma profonde reconnaissance et mes sincères remerciements, de m’avoir fait
l’honneur d’être examinatrice et de participer au jury de ce travail .
Il m’est aussi agréable de remercier Madame Chahrazed
BEKHECHI, maître de
conférences à l’Université de Tlemcen, qu’elle trouve ici l’expression de ma gratitude pour
l’attention qu’elle a bien voulu porter à ce travail, en acceptant de le juger en qualité de
membre de jury.
J’exprime mes vifs remerciements également à tous les membres du Laboratoire des
Travaux Publics de l’Ouest (LTPO) à Tlemcen pour leurs aides dans la réalisation des
analyses pédologiques.
Je tiens à remercier très vivement tous les membres de l’unité « Molécules de
Communication et Adaptation des Microorganismes », particulièrement Madame Manon
VANDERVENNET pour son aide absolument indispensable en microbiologie. Mes
remerciements vont également aux personnes avec qui j’ai pu collaboré notamment,
mademoiselle Carine LOMBARD, Monsieur Jean PEDUZZI, Monsieur Christophe
GOULARD et Monsieur Gérard GASTINE. Je tiens à remercier aussi Madame Yanyan
LI pour sa compétence, son dynamisme et son encadrement dans la partie de biologie
moléculaire. Je remercie sincèrement monsieur Idres MASSINISSA pour sa grande
contribution à l’identification des bactéries lactiques.
Mes remerciements vont aussi à l’égard de Monsieur Hamza BOUABDALLAH
Maître assistant à l’Université de Tlemcen, pour m’avoir donnée les coordonnées
géographiques de nos stations d’échantillonnage.
Je remercie mes collègues et amis pour leur soutien et plus précisément Monsieur
GOUZI Hicham et Monsieur MEBITIL Khalil pour leur aide et son amitié.
J’adresse encore mes remerciements à tous les membres du laboratoire de produits
naturels qui contribuent par leur bonne humeur à créer un cadre de travail agréable.
Pour terminer, je dédie ce manuscrit à ma famille, tout particulièrement mes parents
pour leur soutien, leur amour et leur sagesse qui m’ont permis d’aboutir au grade de
Docteur en biologie et de devenir la personne que je suis. Mes pensées vont à mes frères,
mes sœurs, mes oncles pour leur présence de tous les instants, leur sympathie et leur
encouragement qu’ils m’ont apporté.
Un grand merci à ma femme pour sa sympathie chaleureuse, son appui inestimable
et le sourire dans les moments difficiles et à mes fils pour la joie qu’ils nous procurent.

iii
ـــــــــ
!"#$%&'
()*+. () *+,%%-. // -0 10 23)
4 5-623)02.
728 , 9:/;<.2=>?@ AB)(C2 -D=$%#
().
<# &+% E!3 2=F,G/ )%H !"( @6IJ,KLM(2%,
5 , NOF )PB P5 P(O & /.( Q%@ RN ,% NHE S=F TG :@2U J
V2,(-WStreptomyces P 7< ' ..
-0 8H)/XNX12 -()5># 3+" ),"P (+P 5. )( P
:L %" 2%-E! 1+, RN2 QQ,() )(, %2
AB C2 .RN,%Y 3= (API50 CHL) *+" (ARNr 16S) P 822)
5H% $% $X -2 - G/ Z2 :(65%) Enterococcus P (21%) Leuconostoc P
Lactobacillus
),G/(P Lactococcus )G/ (Weissella )G/(.
?@ (-F 7 LX () NH2 $% AB )( 2=F TG/ C2 4
) ("X [U\" )Staphylococcus aureusP Micrococcus luteusP Bacillus cereus
(Listeria monocytogenes
)/ \"[U )Pseudomonas aeruginosa
Escherichia
(coli
C<=L(Candida albicans) /( F$%HE]YN/N <[.
RN,%9 -WQ%@7D=/TTG/8N,Streptomyces sp.
- 6 72A. 2/)^G6/XT5= "2- AB)( 7K_ C2
&+%$X ().
Q)H) )2=)( ABC2( ' Q3. *+" () K =12. 7
)] `2 3+" $% %2 %" .#J23), 5)J- (HE.
ا تا : (P 5F"2 #P P2=FAB)(C2P !"NH2P
().

iv
Résumé
RésuméRésumé
Résumé
La résistance bactérienne aux antibiotiques et l’émergence de nouvelles maladies
infectieuses justifient l’urgence de disposer de nouvelles molécules antimicrobiennes. Les
peptides antimicrobiens sont des molécules clés dans la défense des organismes vivants face
aux agressions extérieures et aux compétiteurs.
L’objectif de ce travail s’inscrit dans le cadre de la recherche et le criblage des
actinomycètes et des bactéries lactiques capables de synthétiser des substances
antimicrobiennes.
Pour cela, cent huit souches d’actinomycètes ont été isolées à partir d’échantillons de sol
prélevés de différents sites de Tlemcen et Laghouat (forêts, oueds, oasis, montagnes
et barrage). L’identification moléculaire basée sur le séquençage du gène codant l’ARNr 16S
des quarante-sept souches les plus actives de différentes origines, a permis de les rattacher au
genre Streptomyces, souches réputées être la source la plus importante d’antibiotiques.
Aussi dans le cadre de l’exploitation de la diversité microbienne de préparations laitières
traditionnelles algériennes (Jben, Zebda, Dhan et klila), un total de 8 échantillons provenant
de différentes fermes et régions ont fait l’objet d’une analyse microbiologique pour
déterminer leur diversité en bactéries lactiques. Par des méthodes biochimiques (API50 CHL)
et moléculaires (ARNr 16S), on a pu identifier 20 souches qui appartient aux genres
Enterococcus (65%), Leuconostoc (21%), Lactobacillus (2 souches), Lactococcus (1 souche)
et Weissella (1 souche).
Après un criblage initial, le test d’activité antimicrobienne a été effectué sur des souches
d’actinomycètes et de bactéries lactiques contre des bactéries à coloration de Gram positive
(Staphylococcus aureus, Micrococcus luteus, Bacillus cereus et Listeria monocytogenes), des
bactéries à coloration de Gram négative (Pseudomonas aeruginosa, Escherichia coli) et une
levure (Candida albicans) par des méthodes de diffusion sur un agar.
Les souches actives ont montré une forte activité inhibitrice vis-à-vis des
microorganismes pathogènes et les diamètres des zones d’inhibition varient entre 06 et 29
mm pour les actinomycètes et entre 03 et 37 mm pour les bactéries lactiques.
La rechercher par PCR des systèmes génétiques de biosynthèse des peptides lasso par des
Streptomyces sp. n’a pas donné d’amplification d’ADN. Ainsi que des essais d’extraction de
bactériocines produites par des bactéries lactiques n’ont pas abouti à l’isolement de peptides
antimicrobiens.
Les bactéries (actinomycètes et bactéries lactiques) sont une source prodigieuse de
molécules antimicrobiennes d’une grande diversité. Les écosystèmes Algériens abritent une
variété intéressante de microorganismes qui peuvent avoir des activités biologiques.
Mots clés : Sol, Produis laitiers traditionnels, Actinomycètes, Bactéries lactiques, Activité
antimicrobienne, Peptides antimicrobiens.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
1
/
209
100%