Le Cycle cellulaire

PAES 2010-2011
STAGE DE PRÉ RENTRÉE UE 2

Introduction
•Théorie cellulaire : toute cellule est issue
d’une cellule
•L’information génétique est transmise aux 2
cellules filles à chaque division
•Le génome de la cellule eucaryote :
–le matériel génétique est enfermé dans un
compartiment membranaire : le noyau
–matériel génétique = chromosomes

Rappel sur les chromosomes
•Chez l’homme : 23 paires soit 46 chromosomes
contenant chacun 1 molécule d’ADN
•Ne pas confondre brins d’ADN et molécule d’ADN !
1 chromatide = 1 molécule d’ADN = 2 brins d’ADN
1 molécule d’ADN = 1 fibre nucléosomique

•1 chromosome peut contenir 1 ou
2 chromatide(s) selon le moment
du cycle cellulaire
•1 chromatide est formée d’1
molécule d’ADN
•1 molécule d’ADN est formée de 2
brins complémentaires
2 brins d’ADN =
1 molécule d’ADN =
1 chromatide 2 chromatides

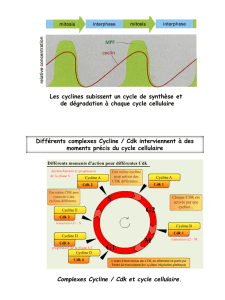
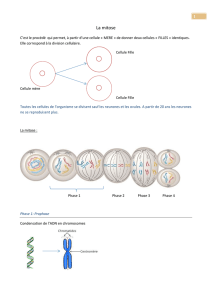
Vue d’ensemble du cycle cellulaire
•Il dure environ 10h chez l’Homme
•La mitose dure environ 1h chez l’Homme
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
1
/
23
100%