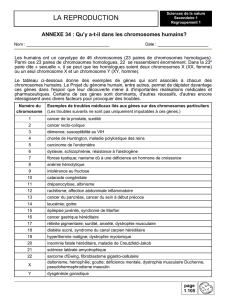
14 la genetique

10/08/2012
1
1
Dr BOGGIO
La génétique
2.2 S1 Cycles de la vie et grandes fonctions
IFSI Dijon - Promotion COLLIERE 2014-2015
14
Les chromosomes sont constitués d’ADN.
Les gènes sont des segments d’ADN.
Ils renferment des recettes, des plans pour la synthèse des protéines.
Une grande partie des protéines sont des enzymes.
Ces enzymes dirigent les synthèses
de presque toutes les molécules de l’organisme.
Par l’intermédiaire des enzymes,
les gènes s’expriment dans la couleur des yeux,
le sexe du sujet, son groupe sanguin,
la proportion des différents types de fibres musculaires…
La génétique est la science de l’hérédité.
L’ensemble des gènes constitue le génome (humain).
Il a été récemment déchiffré :
la séquence des trois milliards des bases de l’ADN
(code génétique) humain est connue.
La fonction de plusieurs milliers de gènes est connue.
Espoir thérapeutique.
Notions de base.
L’être humain possède deux jeux de chromosomes,
un provenant de chaque parent
(figure 16.4)
La plupart des cellules de l’organisme
renferment ce double jeu de chromosomes.
On les dit diploïdes
et le nombre des chromosomes est représenté par 2n.
Chez l’homme : 2 n = 46.
Les cellules diploïdes contiennent 23 paires de chromosomes
dits chromosomes homologues.
Chaque paire est constituée
d’un chromosome paternel et d’un chromosome maternel.
Les deux chromosomes d’une même paire sont semblables.
Ils portent des gènes qui codent pour les mêmes traits
mais pas nécessairement
pour la même expression de ces traits.
Exemple : des gènes homologues
(situés au même endroit sur les chromosomes homologues)
déterminent l’expression des taches de rousseur.
Le gène porté par le chromosome paternel
peut coder pour la présence

10/08/2012
2
2
d’un grand nombre de taches de rousseur,
alors que le gène porté par le chromosome maternel
peut coder pour leur absence.
Les gamètes, cellules sexuelles, renferment seulement
23 chromosomes
soit un nombre haploïde dit n
une seul chromosome
de chaque paire de chromosomes homologues
Quand le gamète mâle et le gamète femelle s’unissent
(fécondation), l’œuf (= zygote) est diploïde.
La formation des gamètes chez l’homme et la femme,
fait intervenir une étape essentielle,
une division cellulaire particulière, la méiose.
Dans la division cellulaire classique (mitose),
les chromosomes répliqués
sont distribués également aux deux cellules filles
(celles qui sont issues de la division).
Chacune des cellules filles reçoit
un jeu de chromosomes strictement identique
à celui de la cellule qui s'est divisée
(la cellule mère).
Chaque cellule issue fille reçoit donc
les deux chromosomes homologues d’une même paire.
La méiose comporte deux divisions cellulaires successives
qui produisent quatre cellules filles.
Chacune de ces cellules filles possède
la moitié des chromosomes de la cellule ordinaire,
un de chaque paire de chromosomes homologues.
La méiose réduit donc le nombre de chromosomes de moitié :
de 2n à n.
Figure 17.1
La première division de la méiose est appelée méiose I
La seconde division de la méiose est appelée méiose II.
Méiose I
Rappel de la mitose :
Avant la mitose, tous les chromosomes sont répliqués
(pendant la fin de l’interphase).
Les copies identiques restent unies
sous forme de chromatides-soeurs
unies par un centromère.
Les centromères se divisent
et les chromatides se séparent
pour migrer chacune vers un pôle de la cellule.
Chaque cellule-fille

10/08/2012
3
3
hérite donc d’une copie de chaque chromosome.
Dans la méiose I,
les chromosomes se répliquent.
Mais les chromosomes répliqués de la même paire se cherchent, se trouvent et
s’apparient sur toute leur longueur.
Ils s’accolent en plusieurs points comme une boutonnière.
Ce phénomène est la synapsis.
Il conduit à la formation de tétrades
(deux chromosomes répliqués de la même paire et appariés).
La synapsis permet l’enjambement ou crossing-over
17.4
L’enjambement est le croisement d’une chromatide maternelle
et d’une chromatide paternelle.
L’enjambement permet l’échange de matériel génétique,
entre les chromosomes paternel et maternel.
Il autorise donc un certain brassage du matériel génétique.
Retour 17.1
Pendant la méiose I,
les tétrades s’alignent au hasard
à l'équateur du fuseau mitotique,
On peut donc trouver des chromosomes maternels et paternels
de chaque côté de la plaque
(le schéma 17.1 ne le montre malheureusement pas !)
Puis les chromosomes homologues d’une même paire
migrent chacun vers un pôle,
mais les chromatides d’un même chromosome
ne se séparent pas.
La méiose I est dite division réductionnelle de la méiose,
puisque les cellules filles sont haploïdes.
(Les chromatides-sœurs unies par le centromère
sont un seul chromosome déjà répliqué).
Méiose II.
C’est ‘équivalent d’une mitose
sauf que les chromosomes ne se répliquent pas
à la fin de l’interphase puisqu’ils le sont déjà ! répliqués.
Les chromatides se répartissent entre les deux cellules- filles.
La méiose II est dite division équationnelle de la méiose.
Résultat : quatre cellules haploïdes,
chacune possédant une seule copie
de chaque molécule d’ADN
La méiose a donc deux effets.
Elle divise le nombre des chromosomes par 2.
Elle crée des variations génétiques.
Le vocabulaire de la génétique

10/08/2012
4
4
46 chromosomes, 23 paires ;
Une des paires est constituée des chromosomes dits sexuels (X et Y) parce qu’ils
déterminent le sexe génétique de l’individu
XX = féminin
XY = masculin
Les 44 chromosomes des 22 autres paires sont les autosomes. Ils guident
l’expression des autres traits.
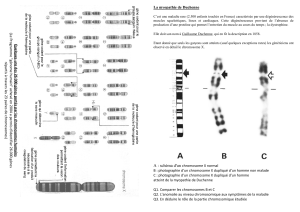
Le caryotype est l’ensemble des chromosomes
d’une cellule diploïde
(17.2)
C’est un examen effectué par les laboratoires de cytogénétique.
Les chromosomes homologues sont classés selon leur taille,
la position de leur centromère,
la longueur des « bras » de part et d’autre du centromère,
les motifs des bandes claires et sombres qu’ils portent.
Un caryotype peut mettre en évidence
une anomalie du nombre ou de la structure des chromosomes.
Le génome est l’ensemble du matériel génétique.
Il est constitué de l’ensemble des molécules d’ADN,
organisé en deux ensembles d’instructions génétiques,
un qui provient de la mère, un qui provient du père.
Le génome est constitué de 25 000 gènes,
inégalement répartis sur les chromosomes.
Chaque gène est une séquence d’ADN
qui donne les instructions nécessaires
à la synthèse d’une protéine.
Un gène a un emplacement précis appelé locus
sur un chromosome.
Exemple le gène qui détermine le groupe sanguin ABO
a le locus 9q34 :
il occupe la quatrième bande de la région 3
du bras long (q) du chromosome 9.
Allèles
Les chromosomes étant appariés, les gènes le sont aussi.
C’est en réalité une paire de gènes
(et non un gène comme dit précédemment)
qui donne les instructions
pour la synthèse d’une protéine
conduisant à un trait particulier.
Les gènes appariés qui occupent
le même locus de chromosomes homologues
sont appelés allèles.
Ils peuvent être exactement identiques
ou présenter des variantes discrètes
dans la séquence des bases.
Dans le premier cas les deux allèles codent
pour la même expression d’un trait,

10/08/2012
5
5
dans le second cas
pour des expressions différentes.
Lorsque les deux allèles qui déterminent un trait
sont identiques,
la personne est dite homozygote pour ce trait.
Lorsqu’ils sont différents, elle est dite hétérozygote.
Parfois un allèle masque ou supprime l’expression de l’autre.
On dit qu’il est dominant.
L’autre est récessif.
L’allèle dominant est représenté par une majuscule,
le récessif par une minuscule.
Les allèles dominants s’expriment,
qu’il y en ait ou deux.
La combinaison AA ou Aa exprime le trait
comme il est prévu par l’allèle A.
Les allèles récessifs ne s’expriment
que s’ils sont en double exemplaire : aa.
Les traits dominants ne s’expriment pas
nécessairement plus souvent que les traits récessifs
car tout dépend de l’abondance relative de chaque allèle
dans la population.
La combinaison aa pour un gène donné
peut être beaucoup plus fréquente
que les combinaisons AA ou Aa.
Le patrimoine génétique d’une personne est son génotype.
La façon dont il se manifeste est son phénotype.
Exemple : la couleur des yeux fait partie du phénotype.
Elle est déterminée par le génotype.
Les sources sexuelles des variations génétiques
A l’exception des jumeaux vrais,
le génotype de chaque humain est différent de celui des autres.
Les humains diffèrent les uns des autres par 3000 gènes.
Cette variabilité résulte de deux phénomènes :
les deux premiers ont lieu lors de la formation des gamètes :
(1) la ségrégation indépendante
des chromosomes homologues,
(2) l’enjambement des chromosomes homologues,
(1) la ségrégation indépendante des chromosomes
17.3
Toutes les paires de chromosomes homologues
entrent en synapsis (s’accolent) pendant la méiose I.
Le synapsis se produit au cours
de la formation des spermatozoïdes,
les gamètes mâles (= spermatogenèse)
 6
7
8
9
10
11
6
7
8
9
10
11
1
/
11
100%