GLUCOSE

Cours de Biochimie PCEM1
année 2004-2005, deuxième semestre
Faculté de Médecine Pitié-Salpétrière, Pr P Ferré
Faculté de Médecine Saint-Antoine, Pr J Capeau
Module Métabolisme glucido-lipidique et régulation
Métabolisme glucidique
Métabolisme lipidique
Régulation du métabolisme énergétique
1

METABOLISME GLUCIDIQUE
•I - Vue d’ensemble
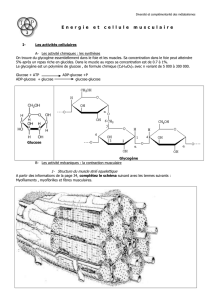
•II - Le transport transmembranaire du glucose
•III - Métabolisme du glycogène
1. Synthèse du glycogène
2. Dégradation du glycogène
3. Utilisation
4. Régulation
•IV - Néoglucogénèse
1. Vue générale
2. Les étapes spécifiques
3. Les substrats
4. Régulation
•V - Voie des pentoses-phosphates
1.Vue générale
2. Deux phases : oxidative et non-oxidative
3. L’utilisation de la voie des pentoses
•VI - Interconversion des oses
1. Métabolisme du fructose
2. Métabolisme du galactose 2

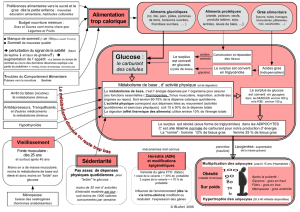
I - Vue d’ensemble
- GLUCOSE
- ACIDES GRAS
Tous les tissus (cerveau : 50% de
l’utilisation journalière).
Muscles, Foie (le cerveau n’oxyde
pas d’acides gras).
Oxydation Mitochondriale
Glucose Pyruvate
ATP
ATP
Glycolyse
Acides gras
3

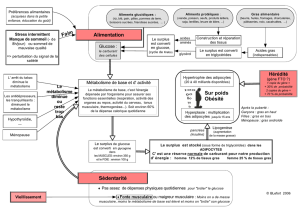
STRATEGIES :
En période d’apport élevé :
- Stocker (glycogène hépatique, lipides)
En période de carence :
-Mobiliser (glycogénolyse)
-Produire de novo (gluconéogenèse)
- Epargner le glucose en mobilisant des substrats de remplacement (lipolyse
libérant des acides gras).
4

5
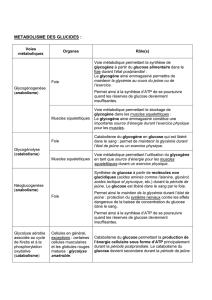
Principales voies du métabolisme glucidique
Milieu extracellulaire
Glucose
Lactate
Cellule
GLYCOGENOGENESE GLYCOGENOLYSE
Glycogène
Glucose 6-Phosphate
GLYCOLYSE NEOGLUCOGENESE
VOIE DES PENTOSES
PHOSPHATE
Lactate Pyruvate
Acétyl-CoA
Mitochondrie CYCLE DE KREBS
PHOSPHORYLATIONS OXIDATIVES
DE LA CHAÎNE RESPIRATOIRE
CO2+ ATP
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
1
/
90
100%