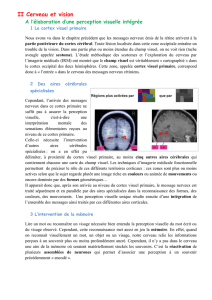
13/03/13 1 Les projections centrales des cellules

13/03/13!
1!
Les projections centrales des cellules ganglionnaires!
Noyau supra-chiasmatique!

13/03/13!
2!
Parenthèse: le cycle circadien veille-sommeil!
Cellules ganglionnaires intrinsèquement photosensibles (mélanopsine) !

13/03/13!
3!
Mélatonine:!
Module les circuits du tronc
cérébral qui contrôlent le
cycle veille-sommeil !
Cellules ganglionnaires M et P!
M (Magnus) :!- grands champs récepteurs!
!!- réponses transitoires à un stimulus soutenu!
!!- suivent rapidement les Δ d’intensité (mouvement)!
P (Parvus) : !- + petites, + nombreuses !
! !- petits champs récepteurs!
!!- sensibles aux Δ de λ (formes et couleurs)!
Cellules ganglionnaires intrinsèquement photosensibles!
(mélanopsine) !
Pas impliquées dans la perception des images!
Rythme circadien, réflexe pupillaire !

13/03/13!
4!
La dégénérescence maculaire
DMLA : Dégénérescence Maculaire Liée à l’Age
DMLA « humide » (10 % des cas)
Formation de nouveaux vaisseaux sous la macula
Progression rapide (pf qq mois)
Traitement laser (photocoagulation)
DMLA sèche (90%)
Disparition épithélium pigmentaire
Perte des photorécepteurs
Pas de traitement
Formes juvéniles: ex maladie de Stargardt
< mutation gène ABC = transporteur de rétinoïdes
Perte progressive des photorécepteurs
-> perte de la vision centrale
Cause la plus fréquente de cécité chez les > 55 ans
Incidence en croissance (vieillissement population)
Diagnostic précoce : grille de Amsler
Causes ??? Hérédité , maladies cardiovasculaires, environnement (tabac, lumière,
nutrition)

13/03/13!
5!
grille de Amsler DMLA débutante
Représentation rétinotopique du champ visuel!
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
1
/
20
100%