Interleukine 6 et myélome multiple chez l`homme - iPubli

Bernard Klein
Régis Bataille
Interleukine
et myélome multiple
chez l'homme
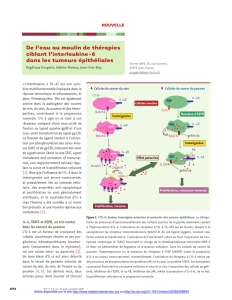
in vivo ex vivo,
In vivo ,
C-reactive protein Ex vivo,

8
h


v
ex vivo.
à

C-reactive protein
Sang périphérique
Moee osseuse
Foie
CRP
 6
7
6
7
1
/
7
100%