Chapitre III Mise en place de la paroi

Chapitre III Mise en place de la paroi :
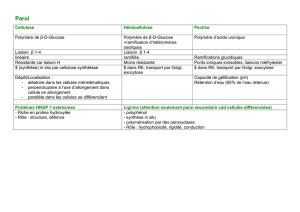
I- Biosynthèse de la cellulose :
1. Glucosyl transférase :
= cellulose synthase (CesA)
Tous les polysaccharides pariétaux sont formés à partir
d’UDP/GDP – D Glucose.
Les enzymes se chargeant des réactions de synthèse de ces
polysaccharides à partir de ce glucose sont des glucosyl
transférases.
Enzyme transmembranaire, elle s’associe en hexamères (6 CesA),
6 hexamères s’associent pour donner des rosettes (=complexe de
36 celluloses synthases -> d’où 36 béta - D – glucoses par
microfibrilles de cellulose).
CesA est une famille multigénique chez les plantes (il en existe des
centaines -> diversité paroi), mais chez toutes les CesA on retrouve
quatre domaines conservés : U1, U2, U3, U4 qui seraient impliqués
dans la liaison du substrat et la catalyse.
2. Origine UDP-glucose ?
a. UGPase :
Enzyme cytoplasmique, transforme glucose 1P en UDP Glucose +
PP avec consommation d’énergie.
b. SuSy :
Enzyme membranaire, transforme sucrose (glucose-fructose) en
UDP glucose + fructose.
3. Le cytosquelette :
Réseau de filaments protéiques (qui s’organisent en câbles) qui
s’étend à travers le cytosol.
Impliqué dans division cellulaire, croissance, différenciation,
déplacement des organites.
Différents types de filaments protéiques :
-> microtubules (tubes) 25nm ;
-> microfilaments d’actines (fins et pleins) 5-7nm ;
-> filaments intermédiaires (on ne connaît pas leur rôle chez
les végétaux)

a. Les microfilaments :
Composés d’actine
Associés dans l’espace avec microtubules et comme ceux-ci, ils
prennent de nouvelles configurations à des moments spécifiques du
cycle cellulaire.
Rôles : courants cytoplasmiques, dépôt de la paroi cellulaire,
croissance terminale des tubes polliniques, déplacement du noyau
avant/après division cellulaire, mouvements des organites et
sécrétion vésiculaire.
Ils interagissent physiquement avec des protéines motrices
(myosines) qui se déplacent le long des microfibrilles -> rôle
notamment dans le mouvement des organelles et le trafic
vésiculaire.
b. Les microtubules :
Dimères de tubuline
alpha et béta,
s’associant en un
protofilament, 13
protofilaments
s’associent en un
microtubule de façon à
former un tube creux
rigide de 25nm.
Ils vont être sans cesse polymérisés/dépolymérisés :
-> forme GTP active
-> forme GDP inactive
Les dimères s’ajoutent toujours à l’extrémité + et doivent être sous
forme GTP pour s’associer mais très rapidement hydrolysé en GDP
instable lorsqu’il est à l’extrémité – et se dissocie -> phénomène de
tapis roulant à l’origine déplacement des microtubules
(polymérisation en + et dépolymérisation en -).
Ils agissent avec différentes protéines :
-> MAP : stabilité des microtubules (empêche catastrophe de
dépolymérisation brutale) par formation de pont entre microtubules.
-> Protéines motrices : rôle dans dynamique des microtubules
(kinésines se déplacent de l’extrémité – vers l’extrémité + et les
dynésines de la + vers la – à la surface des microtubules).
Les microtubules contrôlent l’alignement des microfibrilles de
cellulose qui s’ajoutent à la paroi cellulaire, ces dernières induisant
la direction de l’élongation cellulaire :
Axe microfibrilles transverse à l’axe d’élongation mais parallèle à
l’axe des microtubules.
4. Biosynthèse cellulose :
Soit rosette se retrouve
poussée par microtubule au
cours polymérisation
microtubule avec tapis roulant.
Soit une protéine motrice
interagit avec microtubule et
rosette (cas d’une kinésine
chez une plante).
On ne sait pas encore exactement !

II- Biosynthèse des hémicelluloses et pectines :
(À vérifier)
Synthétisée dans RE
par les ribosomes
Maturation dans
l’appareil de Golgi
(CIS : face RE,
médiane, TRANS :
production vésicules
fusionnant avec
membrane plasmique
pour exocytose) qui
contient les enzymes
nécessaire à cette
étape.
Les pectines :
Elles sont synthétisées (rendues matures ?) par GAUT (enzyme
multigénique) = galacturonosyl transférase.
GAUT 1 et 7 peuvent faire des complexes et sont fortement liées à
la membrane de l’appareil de Golgi côté lumen.
Les pectines peuvent être sous forme méthylées ou déméthylées.
Les PMT (pectine méthyl transférase), exclusivement golgiennes
méthylent pectine dans Golgi.
Les pectines méthylées transitent jusqu’à la paroi.
Les PME (pectine méthyl estérase) déméthylent les pectines ayant
atteint la paroi.
Les pectines déméthylées sont exocytées pour faire le gel
pectinien.
(Ce système de (dé-) méthylation permet de ne pas avoir
constitution du gel à l’intérieur de la cellule et de ne pas modifier le
pH à l’intérieur de la cellule)
III- Structure de la paroi. Les différents types de
liaisons
(Pas complet)
Les fibres de cellulose sont stabilisées par les différents éléments
de la paroi :
- Certaines hémicellulose permettent le positionnement des
fibres cellulosiques entre elles. Par exemple, les
xyloglucanes sont liées à la cellulose par des liaisons
hydrogènes -> mouvement mais aussi stabilité.
- Un réseau supplémentaire de pectines augmente la
complexité de la matrice.
- A la fin de la croissance, le réseau polysaccharidique peut
être fixé par un réseau secondaire de protéines
structurales : comme les HRGP.
Importance des liaisons dans l’architecture de la paroi :
- liaisons hydrogènes : entre fibre de cellulose par exemple ;
- liaisons ioniques : pectines avec Ca++, pectines avec acides
aminés basiques ;
- liaisons covalentes : entre RGII et bore, entre polyosides,
entre protéines (pont isodityrosine et diférulate), entre
polyosides et lignines.

IV- La division cellulaire :
1. Trois mécanismes fondamentaux de la
croissance :
2. Les méristèmes :
Tissu végétal indifférencié dont les cellules sont capables de se
multiplier par mitose.
Assure formation de tissus (histogenèse) et d’organes nouveaux
(organogenèse).
Deux méristèmes : -> apicaux (primaires)
-> caulinaires et racinaires.
Caractéristiques des cellules méristèmales :
- Caractéristiques d’une cellule active : -> forme géométrique
caractéristique : parallélépipède ;
-> rapport nucléoplasmique élevé (=1) : noyau
énorme=toute la cellule ;
-> beaucoup de mitochondries.
- Caractéristiques d’une cellule indifférenciée/totipotente : -> paroi
très mince (primaire), cellules tellement
proches que les virus ne peuvent pas entrer à
l’intérieur de l’organisme ;
-> présence de provacuoles (pas de vraies
vacuoles) et proplastes.
3. La division cellulaire végétale :
La division cellulaire chez les végétaux nécessite de
fabriquer une nouvelle paroi !
Cellule animale : anneau contractile
cellule végétale : dépôt d’une plaque
cellulaire entre deux cellules qui
grandit pour faire la nouvelle paroi.
En interphase les microtubules corticaux sont associés entre eux de
façon similaire aux microfilaments, mais la mitose nécessite une
réorganisation des microtubules : ils prennent la forme de différents
réseaux.

a. Préprophase :
Bande préprophasique = anneau de préprophase.
Elle marque le plan de division de la
cellule et le positionnement du noyau,
c’est à cet endroit que se trouvera la
future paroi entre les cellules filles.
-> structure unique caractéristique de la cellule végétale !
b. Métaphase et anaphase :
Fuseaux mitotiques.
Pendant métaphase : alignement des microtubules sur le plan
équatorial.
Pendant l’anaphase : les microtubules du faisceau mitotique tirent
les chromatides (attachées aux microtubules par leur centromère) -
> séparation des chromatides.
Ces faisceaux sont également présents chez la cellule animale
néanmoins chez les végétaux le pôle des faisceaux mitotiques ne
correspond pas au centriole comme chez les animaux : il y a
plusieurs pôles -> les cellules végétales sont acentriolales.
c. Télophase (fin de télophase -> cytokinèse) :
Pragmoplaste et plaque cellulaire.
Le pragmoplaste permet une accumulation de vésicules (venant de
l’appareil de golgi) au niveau de la plaque équatorial, celles-ci
fusionnent pour donner la plaque cellulaire composée
majoritairement de composés pectiques.
La plaque équatorial s’étend de façon centrifuge jusqu’à atteindre
la paroi cellulaire mère -> séparation totale des deux cellules et
constitution de la nouvelle paroi végétale.
La plaque cellulaire perdure à l’intérieur de la paroi en tant que
lamelle moyenne très riche en pectine qui permet la cohésion entre
les deux cellules.
 6
6
1
/
6
100%