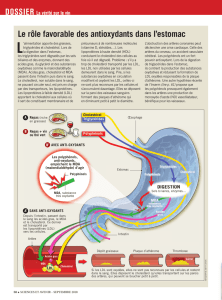
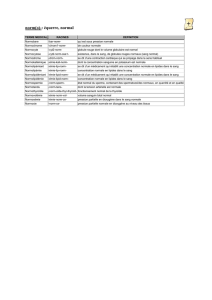
Lipides et lipoprotéines

Université Paris-VI
Lipides et lipoprotéines
Objectifs au cours de
Biochimie PCEM2
Biochimie Métabolique et Régulations
2003 - 2004
Pr. A. Raisonnier ([email protected])
Mise à jour : 16 décembre 2003
Relecture : Pr. A. Raisonnier

2/106 Lipides et lipoprotéines - Pr A. Raisonnier 2003 - 2004

Plan du cours
2003 - 2004 Lipides et lipoprotéines - Pr A. Raisonnier 3/106
Plan du cours
3Plan du cours
7Objectifs
9Partie I : Métabolisme du cholestérol et de ses dérivés
11 Chapitre 1 : Structure du cholestérol
12 1.1 Formule développée du cholestérol
13 Chapitre 2 : Synthèse du cholestérol
14 2.1 Synthèse du cholestérol (définition)
15 2.2 Synthèse du cholestérol (schéma général)
16 2.3 β-céto thiolase (C4)
17 2.4 HMG-CoA synthase
18 2.5 HMG-CoA réductase (enzyme-clé)
19 2.6 Mévalonate kinase
20 2.7 5-PP-mévalonate décarboxylase
21 2.8 Isopentényl-isomérase (I)
22 2.9 Isopentényl-isomérase (II)
23 2.10 Farnésyl-pyrophosphate synthétase (I)
24 2.11 Farnésyl-pyrophosphate synthétase (II)
25 2.12 Coenzyme Q10 = Ubiquinone50
26 2.13 Dolichol-phosphates
27 2.14 Squalène synthétase
28 2.15 Squalène oxydocyclase (I)
29 2.16 Squalène oxydocyclase (II)
30 2.17 Méthyl-hydroxylases
31 2.18 Zymostérol déshydrogénase
32 2.19 Desmostérol réductase
33 2.20 Cholestérol 7-déshydrogénase
34 2.21 Synthèse du cholestérol (schéma général)
35 2.22 Synthèse du cholestérol (bilan) (I)
36 2.23 Synthèse du cholestérol (bilan) (II)
37 2.24 Synthèse du cholestérol (bilan) (III)
38 2.25 Régulation de l’HMG-CoA réductase
39 2.26 Synthèse de l’HMG-CoA réductase
40 2.27 Régulation du taux du cholestérol

Plan du cours
4/106 Lipides et lipoprotéines - Pr A. Raisonnier 2003 - 2004
41 Chapitre 3 : Autres stérols
42 3.1 Synthèse des stérols (schéma général)
43 3.2 Structure du cholécalciférol (vitamine D3)
44 3.3 Synthèse du cholécalciférol
45 3.4 Structure du calcitriol (1,25 diOH-vitamine D3)
46 3.5 7α-hydroxylase
47 3.6 12α-hydroxylase
48 3.7 27-hydroxylase
49 3.8 Structure du cholate
50 3.9 Structure du chénodésoxycholate
51 3.10 Formation des acides biliaires (schéma général)
52 3.11 Cycle entérohépatique des sels biliaires (schéma général)
55 Chapitre 4 : Métabolismes des stéroïdes
56 4.1 Métabolisme des corticostéroïdes (schéma général)
57 4.2 20,22-hydroxylase (I) (enzyme-clé)
58 4.3 Métabolisme des gonadostéroïdes (schéma général)
59 Partie II : Synthèse des lipides
61 Chapitre 5 : Définitions
62 5.1 Synthèse des lipides (définition)
63 5.2 Acylation des lipides membranaires
65 Chapitre 6 : Métabolisme des lipides
66 6.1 Voies de Kennedy (schéma général)
67 6.2 Glycérophosphate acyl transférase
68 6.3 Lysophosphatidate acyl transférase
69 6.4 Phosphatidate phosphatase
70 6.5 Diglycéride acyl transférase
71 6.6 Choline kinase
72 6.7 Cytidine Tri Phosphate = CTP
73 6.8 CDP-choline pyrophosphorylase
74 6.9 Phosphatidyl-choline synthase
75 6.10 Voies de Kennedy (schéma général)
76 6.11 CDP-diglycéride pyrophosphorylase
77 6.12 Phosphatidyl-sérine synthase
78 6.13 Phosphatidyl-sérine décarboxylase
79 6.14 Phosphatidyl-éthanolamine transméthylase
80 6.15 Cycle de la choline (schéma général)

Plan du cours
2003 - 2004 Lipides et lipoprotéines - Pr A. Raisonnier 5/106
82 6.16 Phosphatidyl-inositol synthase
83 6.17 Cycle des phosphoinositides
85 Partie III : Métabolisme des lipoprotéines
87 Chapitre 7 : Nomenclatures
88 7.1 Lipoprotéines du plasma humain
90 7.2 Apolipoprotéines du plasma humain
93 Chapitre 8 : Métabolisme des lipoprotéines plasmatiques
94 8.1 Lipoprotéine lipase
95 8.2 Triglycéride lipase hépatique
96 8.3 Lécithine Cholestérol Acyl Transférase = LCAT
97 8.4 Acyl-CoA Cholestérol Acyl Transférase = ACAT
98 8.5 Récepteur des LDL
99 8.6 Récepteur éboueur
100 8.7 Absorption des lipides
101 8.8 Métabolisme des chylomicrons
102 8.9 Transformation plasmatique des VLDL en LDL
104 8.10 Métabolisme des HDL (I)
106 8.11 Métabolisme des HDL (II)
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
1
/
106
100%