2. Les quatre enzymes spécifiques de la synthèse des acides gras

Biochimie.
Cours d’Inès Masmoudi. PCEM2 2009-2010.
METABOLISME DES ACIDES GRAS
I. Dégradation
- Origine des acides gras :
o Exogène, triglycérides alimentaires : graisses animales, huiles végétales, graines
oléagineuses.
o Endogène, triglycérides du tissu adipeux.
- La dégradation se fait par la β-oxydation : c’est la destruction du squelette carboné en maillons
de 2C.
- Localisation : mitochondries de toutes les cellules sauf les hématies et le cerveau.
- Avant d’entrer dans une voie métabolique, les acides gras voient être activés en Acyl-CoA.
P~P : Pyro-phosphate
1. Passage dans la mitochondrie
- Ce passage nécessite :
o La carnitine, abondante dans le muscule.
o Deux enzymes :
o La CAT (Carnitine-Acyl-Transférase).
o La translocase.

Biochimie.
Cours d’Inès Masmoudi. PCEM2 2009-2010.
2. Différentes étapes
- Le carbone β est oxydé en acide : AcylCoA (n) AcylCoA (n-2)
- Les quatre réactions de la β-oxydation
- Principe : il faut introduire un atome d’oxygène sur le carbone β par addition d’eau. Il faut donc
au préalable créer une double liaison par une déshydrogénase à FAD : on utilise la séquence des
3 réactions qui conduisent au succinate en oaxalo-acétate.

Biochimie.
Cours d’Inès Masmoudi. PCEM2 2009-2010.
- Remarque : Lors de la synthèse des acides gras, on retrouve les réactions de la β-oxydation mais
en sens inverse : les hydrogènes seront fournies par le NADPH.
3. Bilan en énergie
- 18C :
o 9 acétyl-CoA 9x12 = 108.
o 8 FADH2 16 ATP.
o 8 NADH 24ATP.
o Total 148-2 = 146.
- Si on veut comparer le rendement énergétique avec celui du glucose :
o Glucose (6C) 38ATP/6C = 6,3 atomes de carbone.
o Acides gras (18C) 146ATP/18C = 8,1 ATP par carbone.

Biochimie.
Cours d’Inès Masmoudi. PCEM2 2009-2010.
- Les lipides sont plus énergétiques que les glucides. C’est ainsi que notre réserve d’énergie se
trouve sous forme d’acides gras dans le tissu adipeux.
4. Régulation de la β-oxydation
- Chaque cellule a le choix entre deux voies productrices d’énergie : la glycolyse et la β-oxydation.
- Le jeu des régulations hormonales fait qu’en hyperglycémie, après un repas, les cellules oxydent
le glucose. L’excèdent en glucose est transformé en acides gras pour être stocké sous un volume
minimum (sous l’effet de l’insuline, l’hormone de réserve).
- A l’inverse en hypoglycémie, lors du jeûne, il y a sécrétion de glucagon et d’adrénaline. Les
cellules oxydent alors les acides gras et le glucose est épargné pour le cerveau et les hématies.
- La régulation de la β-oxydation se fait au niveau du passage de la membrane mitochondriale :
- La synthèse des acides gras inhibe ainsi leur dégradation : synthèse et dégradation sont deux
voies inverses et ne peuvent coexister.
- Dans la mitochondrie, la β-oxydation est régulée passivement par les rapports FAD/FADH2 et
NAD/NADH au niveau des déshydrogénases. Ces rapports dépendent de la vitesse de la chaine
respiratoire, donc du rapport ADP/ATP : la β-oxydation dépend donc finalement du besoin en
énergie.
- C’est ainsi que l’exercice musculaire, qui consomme de l’ATP ? active la β-oxydation et mobilise
les triglycérides du tissu adipeux.
Maladie génétiques
- Les muscles oxydent préférentiellement les acides gras pour la production d’énergie.
- Il existe des maladies génétiques avec déficit d’enzymes de la β-oxydation, ce qui entraine une
accumulation de lipides dans les muscles :
o Déficit en CAT : Cartnitine Acyl Transférase.
o Déficit en Flavine déshydrogénase (Déshydrogénase I).
On a alors une myopathie (faiblesse musculaire). Diagnostique par biopsie musculaire.
5. destinées de l’acétyl-COA
a. Cycle de Krebs principalement
- L’entrée dans le cycle nécessite de l’oxalo-acétate (OA) qui provient essentiellement du glucose
par le pyruvate : on dit que les lipides brulent au feu des glucides. L faut un peu de glucose pour
que la dégradation des acides gras soit totale.
b. Cétogénèse, si l’oxalo-acétate manque
- C’est le cas :
o Dans le jeûne où l’OA sort de la mitochondrie pour la gluconéogenèse.
o Dans le diabète où le glucose ne peut pas pénétrer dans les cellules : le coma diabétique
est provoqué par l’acidification du sang par les corps cétoniques : c’est l’acidocétose.
- Les trois corps cétoniques sont synthétisés dans le foie :

Biochimie.
Cours d’Inès Masmoudi. PCEM2 2009-2010.
Le β-HMG-CoA (β-Hydroxy-Méthyl-Glutaryl) est un intermédiaire de synthèse du cholestérol.
Destinée des corps cétoniques :
- Synthétisés dans le jeune par le foie, ils constituent une forme de transport des Acétyl-CoA : ils
sont exportés vers les autres tissus, y compris le cerveau, où ils vont redonner des Acétyl-CoA qui
seront oxydés pour donner de l’énergie. Ce sont des combustibles de secours lors d’un déficit en
glucose.
- En cas de surproduction, une partie sera éliminée par les reins, et l’acétone volatile sera
également excrété par les poumons (d’où l’halène des diabétiques).
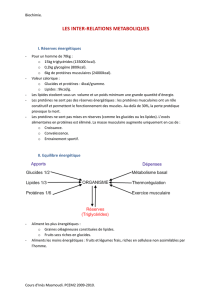
II. Biosynthèse
1. Différentes étapes
- Elle se fait par addition de maillons de 2C (provenant des Acétyl-CoA de la glycolyse), ce qui
explique que les acides gras soient à nombre pair de carbone.
- Elle se fait à partir de l’excédent de glucose de l’alimentation.
- Elle est localisée dans le cytoplasme du foie et du tissu adipeux (alors que la β-oxydation est
mitochondrie).
2. Différentes étapes
- Les intermédiaires sont les mêmes que ceux de la β-oxydation.
- Le coenzyme donneur d’hydrogène est le NADPH (alors que la β-oxydation produit NADH et
FADH2).
- Les enzymes sont associés dans un complexe multienzymatique : la palmitoyl-CoA-synthase, qui
possède une structure comparable au CoA appelée ACP (Acyl Carrier Protein).
 6
7
8
9
6
7
8
9
1
/
9
100%