Régulation hormonale à court terme de la glycémie chez le

Régulation hormonale à
court terme de la
glycémie chez le Rat,
mise en évidence par
l’action de l’insuline
Compte rendu de TP
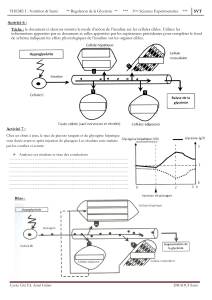
En injectant de l’insuline, hormone pancréatique naturelle,
à un rat vivant anesthésié par un cathéter, on induit
rapidement une hypoglycémie, qui est suivi d’une
augmentation de la glycémie jusqu’à sa valeur initiale. On
met ici en évidence l’action antagoniste des deux hormones
pancréatiques que sont l’insuline et le glucagon sur la
régulation de la glycémie (l’effet hypoglycémiant de
l’insuline et l’effet hyperglycémiant du glucagon). On peut
alors apprécier l’implication de ces deux hormones selon
des conditions physiologiques particulières que sont la
période postprandiale ou une période de jeûne.
Jean SIMONNET
Aurélien CHATEIGNER

2
1 Introduction
La glycémie est la concentration de glucose sanguin. Elle varie en fonction de différents paramètres
que sont notamment l’alimentation ou l’effort physique. On constate que, chez l’homme, la glycémie
normale se situe autour de 1 g/L, et que cette valeur fluctue relativement peu lors d’une journée,
alors que les apports glucidiques ne sont pas les mêmes. Après un repas, le glucose est absorbé au
niveau du tractus digestif et passe alors dans le sang. A distance des repas, il n’y a plus d’apport
glucidique par l’alimentation. Une régulation du taux de glucose sanguin s’effectue et permet un
maintient autour d’une valeur fixe.
On sait que cette régulation met en jeu des hormones, et particulièrement l’insuline, dont le rôle va
être de diminuer la glycémie lorsque celle-ci est trop élevée.
Après injection de cette hormone à un rat, on va observer les fluctuations de la glycémie et alors
déduire les mécanismes physiologiques qui entrent en jeu. Comment et pourquoi l’insuline
engendre-t-elle une diminution du taux de glucose sanguin ? Quels sont les mécanismes cellulaires et
moléculaires qui entrent en jeu ? On déduira alors l’importance biologique d’une telle hormone.
2 Matériel et méthode
On utilise des rats adultes, mâles ou femelles. Ils sont anesthésiés par injection intra-péritonéale
d’uréthane, un anesthésique couramment utilisé, du fait de son coût peu élevé. C’est un bon
anesthésique qui permet une narcose longue (12 heures en théorie), ce qui est très bien pour les
manipulations qui ne vont pas durer plus de 3 heures. En fin de manipulation, une dose forte de ce
même anesthésique est utilisée pour l’euthanasie de l’animal, étant donné que le réveil de l’animal
n’est pas souhaité pour l’expérience. Cependant, il peut occasionner des détresses respiratoires, il
faut donc être vigilant pour pourvoir agir vite en cas de problème. Il cause aussi une légère
hypothermie, on veille à ce que le rat soit placé assez près de la lampe, ce qui permet de la
réchauffer légèrement et de compenser cette perte thermique.
On utilise des techniques chirurgicales : le cathétérisme de la jugulaire et celui de la carotide.
Le cathéter de la jugulaire permet différentes injections. C’est en effet par là que l’on injecte, pour
des raisons anatomiques : la jugulaire est la veine qui apporte le sang directement au cœur. En
injectant ici, on est sûr que le produit sera rapidement diffusé au travers de tout l’organisme. Dans
un premier temps, l’héparine sera injectée. C’est un anticoagulant puissant, elle inhibe un des
facteurs de coagulation. Ceci défavorise la formation de caillots dans les cathéters et empêche le
sang, que l’on prélèvera par la suite, de coaguler, et donc facilite les manipulations, pas toujours
évidentes. On injectera ensuite l’insuline par ce même cathéter, à raison de 0,2 UI pour 100g de
poids d’animal. Notre rat pesant 520g, et la solution étant à 1 UI / mL, on injecte 1,1 mL de solution
par la jugulaire. On défini le temps t0 à partir de cet instant.
On aura préalablement effectué un prélèvement sanguin par le cathéter carotidien, qui constitue
alors le prélèvement t0. On utilise ce cathéter pour le prélèvement car la carotide est l’artère qui fait
sortir le sang directement du cœur, et donc la pression y est importante, ce qui permet un bon débit
sanguin pour le prélèvement. On réalise en tout 6 prélèvements : à t = 0, 10, 25, 40, 60 et 75 min. Le
prélèvement à t0 sert à définir la valeur normale de référence de la glycémie chez le rat.

3
Directement après avoir prélevé, on dilue au 10ème l’échantillon sanguin, pour éviter toute
coagulation ou hémolyse. Pour ce faire, on prélève 100µL de sang que l’on place dans 900µl de
sérum physiologique. On centrifuge ensuite (3000 t/min à 15°C pendant 5min) pour récupérer un
surnageant de plasma sanguin (ici c’est du plasma et non du sérum car c’est du sang non coagulé)
dilué 10 fois. C’est dans ce surnageant que le glucose sanguin est présent. On va l’utiliser pour doser
le glucose par la méthode de Trinder. On réalise également ce dosage pour une solution de liquide
physiologique ainsi que pour une solution de glucose à 1g/L dilué 10 fois (pour que la valeur
d’absorbance soit située dans un intervalle lisible). On lit ensuite les absorbances au
spectrophotomètre à 505nm. On pourra déduire la concentration en glucose de chaque échantillon
par comparaison avec l’absorbance de la solution de glucose à 1 g/L. Les dilutions étant identiques, la
valeur de la glycémie est donnée par le simple rapport DOéchantillon/ DOstandard.
3 Résultats
Temps (min)
0
10
15
40
60
75
Rat 1
2,07
1,82
1,43
1,02
1,39
Rat 2
2,46
2,77
2,38
2,09
2,47
3,1
Rat 3
2,41
2,44
2,34
1,84
2,26
2,51
Rat 4
1,2
0,56
0,67
1,003
1,24
0,95
Rat 5
1,53
1,27
1,24
1,14
1,34
Rat 6
2,78
2,19
1,8
1,84
1,63
1,83
Rat 7
2,76
2,4
2,46
1,07
0,64
0,42
Rat 8
0,87
1,27
0,7
0,78
0,51
Rat 9
1,09
0,36
0,34
0,36
0,36
Rat 10
0,82
1,15
1,21
1,06
0,72
Moyenne
1,80
1,83
1,52
1,21
1,28
1,38
Ecart type
0,78
0,77
0,72
0,55
0,69
1,11
Tableau 1 : Résultats de la glycémie pour chaque rat en fonction du temps. Les valeurs sont données en g/L. Les résultats
personnels sont en italique
Globalement la glycémie chute jusqu’à 40 min, puis remonte ensuite vers la valeur de la glycémie
normale (celle mesuré à t0).
On constate des résultats étranges dont les valeurs seront exclues de l’étude :
Rat 7 : la valeur de glycémie ne fait que chuter.
Rat 8 : les valeurs ne varient pas logiquement (courbe en dents de scies).
Rat 9 : La glycémie chute et reste constante.
Rat 10 : Les résultats oscillent autour d’une valeur moyenne de 0,99 g. A noter qu’une légère
hémolyse s’est produite, ce qui a causé une erreur dans la mesure d’absorbance.
Chaque résultat sera expliqué dans la discussion

4
Temps (min)
0
10
15
40
60
75
Rat 1
2,07
1,82
1,43
1,02
1,39
Rat 2
2,46
2,38
2,09
2,47
3,1
Rat 3
2,41
2,44
2,34
1,84
2,26
2,51
Rat 4
1,2
0,56
0,67
1,003
1,24
0,95
Rat 5
1,53
1,27
1,24
1,14
1,34
Rat 6
2,78
2,19
1,8
1,84
1,63
1,83
Rat 7
Valeurs exclues
Rat 8
Rat 9
Rat 10
Moyenne
2,17
1,92
1,76
1,43
1,57
1,76
Ecart type
0,61
0,77
0,68
0,47
0,63
1,10
Tableau 2 : Tableau des résultats, dans lequel ne sont pas présentes les valeurs exclues.
On déduit une valeur moyenne de la glycémie : 2,17 ± 0,61 g / L.
Pour résumer, on constate une diminution de la glycémie après injection de l’insuline, puis une
augmentation par la suite.
0
0.5
1
1.5
2
2.5
3
3.5
010 20 30 40 50 60 70 80
Glycémie (g/L)
Temps après injection (min)
Evolution de la glycémie en fonction du temps écoulé
après l'injection d'insuline
Rat 1
Rat 2
Rat 3
Rat 4
Rat 5
Rat 6
Rat 7
Moyenne

5
4 Discussion
L’insuline étant une hormone hypoglycémiante, l’effet observé au début, c'est-à-dire la diminution
du taux de glucose dans le sang, est probablement liée à son action. On constate cependant que la
concentration de glucose augmente pour revenir autour de la valeur initiale par la suite, l’organisme
répond à l’hypoglycémie engendrée par l’insuline et évite ainsi l’atteinte d’une valeur trop basse de
la glycémie.
Lors de l’injection, l’insuline, qui est une hormone peptidique, normalement synthétisée par le
pancréas, va induire une diminution de la concentration de glucose sanguin, en influençant d’une
part l’entrée du glucose dans les cellules de l’organisme, et d’autre part sa prise en charge cellulaire
après son entrée dans la cellule, c'est-à-dire en influençant la voie dans laquelle le glucose est utilisé.
L’insuline influence alors différents métabolismes, en fonction du tissu impliqué, chaque tissu tirant
son énergie ou la stockant de différentes manières. Elle peut alors influencer le métabolisme
glucidique de base notamment dans les cellules qui comportent les enzymes permettant le
métabolisme du glycogène ou encore l’équilibre entre glycolyse et néoglucogenèse (hépatocytes,
myocytes…). L’insuline joue également un rôle dans le métabolisme lipidique, notamment en
influençant et en permettant la synthèse de triglycérides et de cholestérol (foie, tissu adipeux).
Dans un contexte normal, ceci se produit juste après un repas riche en glucides où les sucres passent
du tractus digestif vers le sang, ce qui provoque une hyperglycémie, et où le corps à donc la nécessité
de faire baisser cette valeur.
Figure 1 : Molécule d'insuline :
composé de 2 chaines peptidiques
liées par des ponts disulfures
 6
7
8
9
10
6
7
8
9
10
1
/
10
100%