Corrigé Bac blanc 2010

Corrigé Bac blanc 2010
Exercice II 1 Subduction dans la région de Sumatra
La région de l’île de Sumatra est une frontière de plaques tectoniques. Les informations apportées par le document vont
permettre de vérifier que c’est une zone de convergence et de déterminer le sens de cette subduction. /0.25
Les isobathes de la carte fournie indiquent l’existence d’un important relief négatif, la fosse océanique de Java qui desecnd
jusqu’à plus de 7000m. Elle borde l’île de Sumatra parallèlement à un relief positif portant des volcans actifs entourés de
laves et de projections volcaniques qui indiquent un volcanisme de type explosif. On observe aussi le long de la fosse un
chapelet d’îles séparé de Sumatra par le bassin de Montuvaï. La carte informe que ces îles sont formées de terrains
sédimentaires plissés : elles correspondent à des zones émergées d’un prisme d’accrétion. /1
La région est également caractérisée par une activité sismique intense avec une distribution particulière des foyers des
séismes : ceux-ci sont de plus en plus profonds lorsqu’on s’éloigne de la fosse en direction de l’île, ce jusqu’à plus de 200
km de profondeur. Or les foyers sismiques ne peuvent se trouver que dans la lithosphère cassante dont l’épaisseur moyenne
est de 100 km. Le plan suivant l’emplacement des foyers sismiques, appelé plan de Wadati-Bénioff, matérialise donc
l’enfoncement de la plaque indienne sous la plaque eurasienne portant Sumatra. Une partie des sédiments portés par la
plaque subduite sont « rabotés » et s’empilent sous forme d’écailles en surface, constituant le prisme d’accrétion évoqué ci-
dessus. /1,5
Bilan
L’ensemble des caractéristiques recensées sur les documents sont les marqueurs d’une zone de subduction intra-océanique,
où, ici, la plaque indienne converge vers la plaque eurasienne et s’enfonce sous elle, d’où les séismes mentionnés. /0.5
Faire un schéma très simple, titré et légendé /0.75
Exercice II 2 Obligatoire Homo floresiensis
Des fossiles d’homininés découverts en Indonésie sur l’île de Florès ont conduits leurs découvreurs à créer une nouvelle
espèce. Les documents fournis vont nous permettre de caractériser et de justifier la création d’Homo floresiensis. /0,25
Les données relatives au fossile LB1 et aux restes associés sont rassemblées dans le document 3. Pour pouvoir être rattachés au
rameau humain, ces fossiles doivent présenter au moins un des caractères dérivés propre à l’homme. Le crâne de la femelle
LB1 est arrondi et constitué par une boîte épaisse ; l’arcade zygomatique est large ; les volumes crâniens sont compris entre
380 et 530 cm3 ; le trou occipital semble situé sous la boîte crânienne, indiquant que le fossile est bipède, caractère dérivé
qui fait bien de lui un homininé. La face de LB1 est peu prognathe avec un bourrelet sus orbitaire peu prononcé. On nous
indique également que l’individu, comme ses congénères, était de petite taille : entre 1,06 et 1,20 m. /1,5
Le tableau du document 1 présente en parallèle des caractéristiques d’homininés déjà bien connus : Australopithecus, Homo
erectus et H. sapiens. En les comparant à celles de LB1, on pourra replacer ce dernier dans la classification des homininés.
C’est du crâne d’H. erectus que ressemble le plus celui de LB1 mais il a un volume beaucoup plus faible, de l’ordre de celui
d’Australopithecus. Sa taille est aussi bien plus faible que celle du « grand » H. erectus, 1,60m en moyenne. Cette dernière
donnée peut expliquer le volume crânien, proportionné à la petite taille du sujet. LB1 serait donc en quelque sorte un H. erectus
miniature. /1
Les H. erectus ne sont cependant pas définis que par des caractéristiques physiques : cette espèce avait une culture lithique bien
développée dont l’emblème est le biface, silex taillé par eliminations d’éclats sur ses 2 faces. Les spécialistes pensent qu’il
existait des ateliers de taille au sein des populations d’H. erectus, ce qui sous entend une organisation sociale et la capacité de
langage pour la transmission de cette culture. D’autre part, des traces de foyers sont souvent associées aux restes d’H. erectus,
ce qui montre qu’il maîtrisait le feu. C’est d’ailleurs la première espèce d’Homininés à présenter cette capacité. Rapprocher
LB1 d’H. erectus oblige à vérifier si lui aussi partage ces technique et culture. La seconde partie du document 3 apporte à ce
sujet tous les renseignements nécessaires. Les fouilles qui ont mis à jour LB1 ont permis de trouver des outils en pierre taillée,
variés et spécialisés : pointe, poinçon, lame associés à des restes d’éléphants nains. Les Homo de Florès avaient donc
développé une industrie lithique performante, plus complexe même que celle des H. erectus, qui leur permettait de chasser
ces éléphants ; mais le texte souligne que la chasse aux éléphants nécessite la coopération de plusieurs chasseurs … ce qui
laisse supposer l’existence d’une organisation sociale et d’un langage. Les éléphants tués pouvaient être cuits puisqu’on en
retrouve des restes carbonisés, ce qui indique ces populations de Florès avaient acquis la maîtrise du feu. /1,5
Comme leurs caractéristiques morphologiques, les spécificités techniques et culturelles du petit peuple de Florès le rapprochent
des H. erectus. Il faut maintenant se demander qu’elle pourrait être leur origine.
Le document 2 informe sur la colonisation de la planète par les homininés. On peut constater que des restes d’Homo erectus,
grand voyageur, prouvent sa présence à Java de -1,8Ma à -25000ans. L’île de Florès est, elle, habitée depuis 95000ans. Il est
tout à fait justifié de penser que la colonisation de Florès s’est faite depuis Java par un petit groupe d’H. erectus, qui
aurait emporté avec lui la culture spécifique de cette espèce. L’isolement géographique sur l’île de Florès a pu amener à une
évolution spécifique des individus, un nanisme notamment et le perfectionnement de l’industrie. /1,5
L’étude de ces documents permet donc bien de comprendre pourquoi les paléontologues de Florès ont proposé de classer les
restes des petites hommes de Florès dans une nouvelle espèce, Homo floresiensis, descendant d’H. erectus. Cette espèce
s’est d’ailleurs maintenue sur l’île plus longtemps que les populations d’H. erectus voisines et a vécu contemporainement à
l’installation d’H. sapiens en Australie (-50000ans)et sur Bornéo (-40000ans). /0.5

Corrigé Bac blanc 2010 ROC
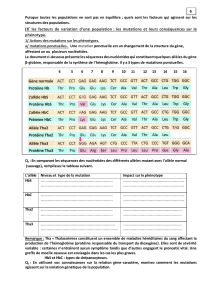
Après avoir exposé la nature des innovations génétiques, expliquez à l’aide d’un exemple précis comment un milieu variable
dans le temps et dans l’espace intervient dans la conservation de ces innovations.
La reproduction permet aux espèces de se conserver au cours des générations successives. Pourtant l’étude des données
paléontologiques accumulées au cours des temps géologiques et les observations du monde vivants ont amené les naturalistes,
comme Darwin, a conclure que les espèces évoluaient, c’est à dire se transformaient, ce en lien souvent avec leur milieu de vie.
Mais quels sont les mécanismes de ces changements ?
Les modifications étant transmises de génération en générations, on sait aujourd’hui qu’elles mettent en jeu les gènes : elles
résultent de ce qu’on peut appeler des innovations génétiques ; les principales seront présentées en première partie de ce
devoir. Nous exposerons ensuite un exemple de conservation d’innovation génétique par la sélection naturelle exercée par le
milieu. /0.5
1. Les innovations génétiques
1.1. Les mutations permettent la création de nouveaux allèles /3
Une mutation est une modification accidentelle de la séquence nucléotidique de la molécule d’ADN.
Une mutation ponctuelle correspond à la modification d’un nucléotide ou de quelques nucléotide de chaque brin d’ADN. Une
substitution est le changement d’un nucléotide, une insertion est l’addition d’un nucléotide et une délétion la perte d’un
nucléotide. Il existe aussi des mutations concernant la délétion de plusieurs milliers de nucléotides.
Les mutations se répartissent au hasard sur l’ADN : l’emplacement d’une mutation est impossible à prévoir. Elles se produisent
spontanément à la suite d’accidents (réplication erronée de l’ADN, mauvaise réparation) et sont donc rares : en moyenne une
erreur lors de la réplication survient avec la fréquence très faible de 1/109 nucléotides soit, vue la taille du génome humain, 3,5
mutations lors de chaque réplication/division mais si on considère l’ensemble des gènes d’un organisme, ou d’une population,
elles peuvent presque être considérées comme fréquentes !
On considère qu’ainsi, dans l’espèce humaine, plus d’1% des gamètes produits sont porteurs d’un nouvel allèle.
Les agents mutagènes (UV, radioactivité, certaines substances chimiques) augmentent leurs fréquences.
Seules les mutations qui touchent les cellules germinales, à l’origine des gamètes, peuvent être transmises à la génération
suivante. Bien que le taux de mutation soit faible, il est suffisant pour faire apparaître de nouveaux allèles par accumulation de
mutations aux cours des générations.
Toutes les mutations n’ont pas la même conséquence sur le phénotype. En effet, la transcription répercute sur la séquence
nucléotidique de l’ARNm une mutation de la séquence codante d’un gène. Le code génétique étant redondant, une substitution
peut ne pas modifier la signification des codons de l’ARNm et être sans conséquence sur la structure primaire de la protéine
codée par le gène muté. La mutation n’a alors pas d’effet sur le phénotype : elle est dite silencieuse.
Une substitution non silencieuse peut entraîner le remplacement d’un acide aminé dans la protéine traduite : c’est une mutation
faux-sens; ce nouvel acide aminé peut provoquer la perte de l’activité biologique ou une activité biologique différente ou
encore ne pas perturber la fonction du polypeptide, la mutation faux sens est alors dite conservatrice (et dans ce cas, il n’y a pas
de conséquences sur le fonctionnement cellulaire, ni sur le phénotype de l’individu). Une substitution peut d’autre part
provoquer l’apparition d’un codon stop prématuré dans la molécule d’ARNm, on parle de mutation non sens, le polypeptide
synthétisé est alors plus court que la protéine de référence et en général non fonctionnel, perturbant le fonctionnement de
l’organisme.
Les insertions et les délétions sont des mutations décalantes qui modifient obligatoirement le cadre de lecture de l’ARNm lors
de la traduction (sauf si elle concerne 3 nucléotides). Elles font très souvent apparaître un codon stop prématuré dans l’ARNm.
La séquence de la protéine est très perturbée, généralement raccourcie et donc non fonctionnelle.
D’autre part, selon le type de gène muté, les conséquences seront différentes sur l’organisme ; certains gènes codent pour des
protéines essentielles au bon fonctionnement de l’organisme, alors que d’autres sont secondaires. Cas le plus lourd de
conséquence est une mutation sur un des gènes du développement qui contrôlent le fonctionnement de toute une cascade
d’autres gènes lors du développement embryonnaire.
Les mutations ont donc des effets variables et certains des nouveaux allèles ainsi créés peuvent être à l’origine d’une
modification du phénotype. Elles sont à l’origine d’une diversification du génome, mais sans en augmenter la taille. Or
l’évolution des êtres vivants s’est accompagnée d’une augmentation spectaculaire de la taille des génomes. Il existe en effet
d’autres mécanismes de modification du génome.
1.2. La duplication de gènes permet la création de nouveaux gènes /1
Les généticiens ont constaté qu’au sein du génome d’une espèce, il existe des gènes différents (à des locus différents) dont les
séquences nucléotidiques présentent des similitudes trop importantes pour que cela soit dû au hasard : ce sont des gènes
apparentés ; leur existence ne s’explique que par la duplication d’un gène ancestral, c’est à dire la copie à l’identique d’un gène
préexistant.
Chaque nouveau gène peut alors être modifié alors par accumulation de mutations. Il y a donc évolution divergente des
duplicata. Un duplicata peut ainsi acquérir une nouvelle fonction au lieu de seconder le gène ancestral dans une même
fonction. Le génome de l’espèce s’enrichit donc progressivement au cours de son évolution et des générations. C’est ainsi que
l’espèce peut acquérir de nouvelles innovations évolutives. Exemples et schéma possible.
(remarque : transposition hors sujet, consigne à respecter)
2. La conservation des innovations génétiques

2.1. la sélection naturelle
Les innovations génétiques étant aléatoires, les individus d’une population sont génétiquement différents.
La plupart des mutations qui se produisent en une génération sont défavorables car elles perturbent le fonctionnement de
protéines indispensables à la vie ; les individus qui la possèdent survivent plus difficilement et se reproduisent moins bien.
Cependant certaines mutations peuvent conférer un avantage dans un milieu donné : si les porteurs d’un allèle muté sont plus
aptes à survivre, à parvenir à la maturité sexuelle, à se reproduire, ils ont une descendance plus nombreuse. Dans ce cas la
fréquence des allèles qu’ils portent augmente dans cette population au fil des générations. Ce mécanisme qui correspond à
l’augmentation ou la diminution rapide au cours des générations de la fréquence d’un allèle qui, dans un milieu donné, favorise
ou défavorise la reproduction sexuée des individus porteurs de cet allèle est appelé sélection naturelle. /1
La sélection naturelle est liée à des conditions environnementales données. La fréquence d’un allèle n’est donc pas la même
dans les populations vivant dans des environnements différents et elle est susceptible de varier si les conditions
environnementales changent.
On peut illustrer ce qui vient d’être décrit par l’exemple du mélanisme de la phalène du bouleau (ou de l’allèle HbS
responsable de la drépanocytose.)
2.2. l’exemple de la phalène du bouleau /2
On connaît pour ce papillon de nuit 2 phénotypes : couleur claire et couleur sombre. Etudié en Angleterre, il n’existait que des
formes claires jusqu’à ce qu’une mutation fasse apparaître la couleur sombre. En simplifiant, on peut dire que ces 2 phénotypes
résultent de l’existence de 2 allèles : S dominant conférant la couleur sombre et c. Mais les formes sombres sont restées rares
jusqu’au XIXème siècle. Cela s’explique parfaitement par la sélection naturelle. En effet, le jour les phalènes se posent sur les
troncs d’arbres, troncs clairs de bouleaux ou troncs d’autres espèces, plutôt clairs aussi à cause des lichens les recouvrant. Dans
ce milieu, les formes sombres étaient donc facilement repérées par les oiseaux, principaux prédateurs. La couleur sombre
conférant un désavantage sélectif, la fréquence de l’allèle S restait très faible et les individus sombres peu nombreux. Mais au
XIXème siècle les entomologistes ont constaté une nette augmentation de la fréquence des formes sombres dans certaines
régions. C’est un changement des pressions sélectives exercées par le milieu qui permet de l’expliquer. Dans ces régions à
cette époque, l’industrialisation a été très forte et la pollution engendrée a noirci les troncs et fait mourir les lichens. Dans ce
contexte pollué, ce sont cette fois les formes claires qui sont défavorisées puisqu’elles sont plus facilement détectables sur les
troncs foncés. Les individus porteurs de l’allèle S, favorisés par leur homochromisme (ndlc : involontaire !!) ont pu mieux
survivre et se reproduire et l’allèle S s’est répandu dans les populations de Phalène des zones polluées. Depuis quelques
décennies la pollution diminue nettement et on voit baisser la proportion des formes sombres. Cet exemple confirme bien que
les mutations qui confèrent un avantage sélectif aux individus qui en sont porteurs ont une probabilité plus grande de se
répandre dans la population.
(ou
S, à l’état homozygote, est très fréquent en Afrique dans les régions ou sévit le paludisme. L’allèle
S à l’état
hétérozygote protège contre le paludisme. La sélection naturelle favorise la reproduction des hétérozygotes par rapport aux
homozygotes (
A//
A) et maintient l’allèle
S à une fréquence élevée.)
(Remarque : la diversité allélique est donc indispensable à la survie d’une espèce. Le polyallélisme seul peut conférer la
capacité de faire face à une modification des contraintes environnementales.
Une sélection diversifiante favorisent l’équilibre entre différents allèles.
une mutation uniformisante tend au contraire à l’élimination d’un allèle.)
Conclusion :
Ainsi, ce sont des évènements rares et aléatoires qui sont à l’origine des innovations génétiques. La sélection naturelle, liée aux
caractéristiques du milieu, en un endroit donné et à un moment donné, permet alors la conservation des innovations les plus
avantageuses pour les individus qui les portent. Les innovations désavantageuses, dans un milieu donné seront éliminées. On
doit également évoquer le cas des innovations génétiques neutres qui ne sont ni avantageuses ni désavantageuses : leur
maintien est aléatoire, elles peuvent se répandre ou tout aussi bien être éliminées ; c’est le phénomène de dérive génétique. /0.5
Le schéma bilan ci-après résume ce sui a été exposé. /1
On peut maintenant se demander par quels mécanismes la conservation de certaines innovations aboutissent à l’apparition de
nouvelles espèces.

Corrigé Bac blanc 2010 Spécialité
Expliquez comment les structures de la feuille coopèrent dans la réalisation de la photosynthèse. (Sujet national
juin 2003)
La photosynthèse est la synthèse de matière organique à partir de carbone minéral (CO2 atmosphérique) grâce à
l’énergie lumineuse. Chez les végétaux chlorophylliens, elle est réalisées par les feuilles. Les documents fournis
vont nous permettre de montrer comment plusieurs types de cellules et d’organites coopèrent pour assurer cette
autotrophie. /0.5 (0.25 définition)
Le document 1 présente les résultats d’une expérience consistant à marquer le CO2 en le rendant radioactif avant
de le fournir à une feuille à moitié éclairée, ce au niveau d’une zone localisée de sa face inférieure.
L’autoradiographie de cette feuille révèle ensuite où le CO2 marqué a été incorporé dans des molécules organiques:
ces zones apparaissent sombres. On constate que la fixation de CO2 d’est faite uniquement dans la zone éclairée : la
lumière est donc indispensable à la synthèse de glucides à partir de CO2 atmosphérique, d’où le terme de
« photo »synthèse. /1
En détaillant la zone d’incorporation, on peut constater que ni les nervures, ni le pétiole ne contiennent de glucides
radioactifs. La coupe de feuille du document 3a laisse penser que la zone sombre d’incorporation de CO2 marqué
correspond aux parenchymes chlorophylliens. /0.5
Le document 2 nous permet de le vérifier. Il donne l’équation de fixation du CO2 sur le ribulose biP qui forme un
intermédiaire non glucidique (l’acide phosphoglycérique) pour la synthèse de glucides. Cette réaction est catalysée
par l’enzyme carboxylase, qui est donc essentielle pour la photosynthèse. Par la technique d’immunofluorescence,
on peut localiser la présence de cette enzyme : sur la coupe de feuille de betterave traitée, en comparant avec la
structure foliaire du document 3, on constate que toutes les cellules des parenchymes chlorophylliens (et
uniquement elles) contiennent l’enzyme. La ribulose biP carboxylase est plus précisément contenue dans les
chloroplastes des parenchymes foliaires. Les pigments des chloroplastes permettent d’autre part de capter l’énergie
lumineuse ; les chloroplastes sont donc le lieu de la photosynthèse, où le CO2 est incorporé dans des molécules de
glucide en présence de lumière et de CO2. /1
Cependant, les parenchymes chlorophylliens ne sont pas en contact avec l’air extérieur. Tout le limbe est recouvert
de cellules épidermiques imperméabilisées en surface par une cuticule. Comment le CO2 atmosphérique arrive-t-il
aux cellules chlorophylliennes ? La coupe transversale du document 3a révèle la présence d’interruption dans
l’épiderme inférieur de la feuille : des stomates sont en effet répartis dans l’épiderme inférieur ; ils sont constitués
par 2 cellules stomatiques délimitant un ostiole, ouverture vers une chambre sous stomatique ; cette dernière est en
relation avec les nombreux méats, espaces entre les cellules du parenchyme lacuneux. Lors de l’expérience du
document 1, le CO2 marqué a été amené au niveau de l’épiderme inférieur : le CO2 radioactif est donc entré dans la
feuille par des stomates puis a diffusé vers l’ensemble de la feuille, passivement, selon les gradients de
concentration ; les cellules chlorophylliennes utilisant ce CO2, la diffusion se fait donc essentiellement vers elles. /1
Le graphe du document 3b indique cependant que les stomates ne sont pas toujours ouverts. Chez l’arbousier, ils
sont fermés pendant la nuit, s’ouvrent de manière importante, jusqu’à 80% de leur ouverture maximale, à partir de
9h du matin jusqu’à 11h ; aux heures les plus chaudes, ils se referment partiellement (à 45%) puis s’ouvrent à
nouveau à partir de 15h pour se refermer au coucher du soleil. La courbe c est celle de l’intensité de la
photosynthèse au cours d’une journée ; elle est très corrélée à celle d’ouverture des stomates. Pendant la nuit il ne
peut y avoir de photosynthèse (pas de lumière) et les stomates sont fermés. Le jour, les maxima d’activité
photosynthétiques (20 et 17 ng deCO2/cm2/s) correspondent aux maxima d’ouverture et inversement, autour de 13h
le minimum d’ouverture des stomates correspond au minimum de fixation du CO2 (moins de 5 ng/cm2/s). Une
bonne fixation du CO2 nécessite donc une bonne ouverture des stomates. La « dépression de midi » est liée à
l’équilibre que doit respecter la plante entre photosynthèse et pertes d’eau, la vapeur d’eau sortant des feuilles par
ces mêmes stomates. /1.5
Toute l’organisation de la feuille est liée à son rôle photosynthétique et ses différentes structures coopèrent :
stomates, parenchyme lacuneux et palissadique. L’ouverture des stomates permet l’entrée de CO2,qui diffuse à
travers le parenchyme lacuneux vers toutes les cellules chlorophylliennes. A la lumière, la fixation de ce CO2 dans
des glucides est réalisées au niveau des nombreux chloroplastes des cellules chlorophylliennes. /0.5
1
/
4
100%