stabilité et variabilité des génomes et évolution

STABILITE ET VARIABILITE DES GENOMES ET EVOLUTION
VOCABULAIRE
Génome : ensemble des gènes
Gène : portion d’ADN codant la synthèse d’une protéine ; c’est une séquence nucléotidique localisée sur
un chromosome en un locus ; il définit un caractère
Génotype : ensemble des allèles
Allèles : différentes versions d’un gène ; il définit l’état d’un caractère
Caryotype : représentation ordonnée des chromosomes d’une cellule
Formule chromosomique : nombre de chromosomes d’une cellule, sous la forme n = … si la cellule est
haploïde, 2n = … si la cellule est diploïde
Haploïde : qui contient un lot de chromosomes
Diploïde : qui contient deux lots de chromosomes homologues
Cellule somatique : cellule de l’organisme autre qu’une cellule germinale
Cellule germinale : cellule à l’origine des gamètes
Dicaryotique : qui contient deux noyaux
L’expression du génome amène la synthèse de protéines de structure et de protéines fonctionnelles
(enzymes, hormones) qui permettent la réalisation des caractères de l’individu. Un gène est dit
polymorphe quand il se présente au moins sous deux formes d’allèles et que ces formes sont présentes
chez au moins 1% des individus. Les individus et les cellules haploïdes possèdent un seul allèle du gène.
Les diploïdes en possèdent deux.
LES INNOVATIONS GENETIQUES
Il y a deux types d’innovations génétiques :
- production de nouveaux allèles et répartition dans la population à l’origine du polymorphisme
génique et phénotypique
- production de nouveaux gènes, ce qui permet l’acquisition de nouvelles fonctions, de nouveaux
caractères
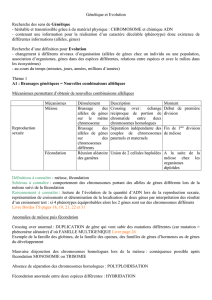
I - Les mutations sont à l’origine du polymorphisme des gènes
- mutation : processus par lequel née un allèle à partir d’un allèle préexistant
A. Les mutations ponctuelles
1) Les mutations ponctuelles non décalantes : les substitutions
a) Les mutations silencieuses
- pas de modification apparente du produit du gène
- premier cas : acide aminé codé par le codon muté identique à celui codé par le codon « normal »
(redondance du code génétique)
- deuxième cas : acide aminé changé (faux-sens) mais pas localisé au niveau du site actif de la protéine
site actif non affecté, donc protéine opérationnelle
b) Les mutations non-sens

- un codon sens ( acide aminé) mute et devient un codon non sens ( stop)
- peptide formé plus court fonction modifiée voir supprimée
c) Les mutations faux-sens
- codon sens remplacé par un autre codon sens ne codant pas pour le même acide aminé
- fonction du peptide modifiée ou non selon que l’acide aminé fasse partie ou non du site fonctionnel de
la protéine
2) Les mutations décalantes : insertion et délétion
- mutations qui entraînent un décalage du code de lecture du ribosome modification profonde de la
séquence d’acides aminés après la mutation
- statistiquement, du fait du hasard, apparition d’un codon stop après une vingtaine d’acides aminés
- fonction protéique généralement profondément changée
3) Bilan
- capacité de la protéine « mutante » à modifier le phénotype cellulaire, voire macroscopique
- mutation d’un gène architecte apparition de nouveaux plans d’organisation mis en place lors du
développement embryonnaire
- action en cascade de ces gènes : mutation d’un gène multiples conséquences
- mutations conservées au niveau des gènes du développement apparition de nouveaux plans
d’organisation nouveaux groupes dans les règnes animal et végétal
B. Expression des allèles au niveau du phénotype
- un gène s’exprime quand la protéine dont il gouverne la synthèse est produite
- individu homozygote pour un gène les 2 allèles gouvernent la synthèse de la même protéine
- individu hétérozygote pour un gène 2 cas possibles :
∟ expression des 2 allèles, chaque protéine définit un trait du phénotype allèles codominants
(groupe sanguin AB)
∟ expression des 2 allèles mais fonction biologique assurée par une des 2 protéines allèles
dominant et récessif (allèles A et O)
- un phénotype macroscopique des génotypes différents
- similitude phénotypique possibilité qu’il y ait des différences cellulaires et moléculaires
C. Origine et transmission des mutations
- mutation : modification aléatoire et accidentelle de la séquence d’ADN, arrivant lors d’une erreur de
réplication ou de réparation de l’ADN
- nature de la mutation indépendante des caractéristiques du milieu
- fréquence des accidents normalement faible (10-6/gène et /gamète chez l’homme)
- nombre de gènes et de nucléotides élevé (3.109) phénomène pas exceptionnel
- augmentation de la fréquence de mutation par des agents mutagènes (UV, rayons X, fumée de cigarette,
amiante…)
II - Les duplications sont à l’origine de nouveaux gènes
- possibilité de duplication d’un gène
- duplicatas transposés à un autre endroit du génome, sur le même chromosome ou pas, association ou
fusion ou non avec des gènes préexistants
- évolution de manière indépendante des duplicatas, pouvant subir des mutations et des duplications
formation de familles multigéniques (différentes chaînes de globines)
- divergence entre gènes d’une même famille accumulation des mutations

- possibilité d’acquisition de nouvelles fonctions (hormones hypophysaires)
- familles multigéniques : témoins de la possibilité de complexification du génome (plasticité du génome)
- besoin des innovations génétiques d’être transmises à la descendance pour jouer un rôle dans l’évolution
III - Conditions de transmission des informations et création de nouveaux génotypes
conditions de conservation d’une innovation dans la population :
- ne pas trop affecter le fonctionnement de la ¢, ne pas entraver le développement et la reproduction
des individus, sinon innovation létale
- affecter les ¢ de la lignée germinale : innovation transmise à toutes les ¢ issues de la ¢ mutée par mitose
∟ si ¢ stomatique touchée, seul le tissu touché manifeste la mutation, pas de transmission à la
descendance
∟ si ¢ de la lignée germinale (à l’origine des gamètes) touchée, mutation transmise à toutes les ¢
de la génération suivante
Les mutations peuvent être pathogènes, indifférentes ou bénéfiques (résistance à une maladie). De toute
façon elles font naître de nouveaux allèles qui viennent enrichir la collection d’allèles de la population. Le
polymorphisme génique actuel semble donc être le résultat des mutations survenues dans le passé au sein
des populations de l’espèce. De plus au cours de l’évolution de nouveaux gènes sont apparus par
duplication, entraînant une complexification du génome. Le moteur des innovations génétiques est donc
purement aléatoire.
Ces innovations peuvent conduire à une individualisation d’une population aboutissant à son isolement
reproductif puis à l’émergence d’une nouvelle espèce (homininés au sein d’une population ancestrale
commune avec celle des chimpanzés). Les phénotypes nouveaux qui en résultent vont être alors
confrontés aux conditions écologiques du milieu dans lequel ils apparaissent. Ainsi, au fil des temps
géologiques des groupes nouveaux sont apparus, se sont diversifiés puis ont régressé et disparu, remplacés
par d’autres, mieux adaptés aux niches écologiques qu’ils occupaient.
MEIOSE ET FECONDATION PARTICIPENT A LA STABILITE DE L’ESPECE
La reproduction sexuée est le processus qui permet la production d’un nouvel individu à partir de 2
individus (animaux ou végétaux) de même espèce et de sexe différent. On appelle cycle de développement
ou cycle biologique la succession des diverses phases d’une génération qui se déroule depuis l’œuf jusqu’à
la production d’un nouvel œuf.
I - La méiose permet le passage d’un état diploïde à un état haploïde
A. Principales étapes
- prophase I : condensation des chromosomes à 2 chromatides dans le noyau, disparition de la membrane
nucléaire, crossing-over
- métaphase I : alignement des chromosomes homologues sur la plaque équatoriale
- anaphase I : séparation des chromosomes homologues, chaque chromosome de chaque paire part dans
la direction opposée, apparition d’un sillon de division
- télophase I : division des 2 ¢, passage d’une ¢ à diploïde à 2 ¢ haploïdes
- prophase II : 2 ¢, chromosomes condensés
- métaphase II : alignement des chromosomes sur la plaque équatoriale
- anaphase II : séparation des chromatides

- télophase II : division des ¢, formation de 4 haploïdes avec des chromosomes à une chromatide
- étapes télophase I et prophase II confondues, les 2 divisions se suivent immédiatement
- méiose I = division réductionnelle
- méiose II = division équationnelle
- différences essentielles avec une mitose : appariement des chromosomes homologues en prophase I
(formation de tétrades de chromatides), séparation des chromosomes homologues et pas des chromatides
en anaphase I
B. Évolution de la quantité d’ADN
- interphase : Q pendant la phase G1, augmentation progressive pendant la phase S jusqu’à 2Q pendant la
phase G2
- méiose : chute à Q en méiose I, puis à Q/2 en méiose II
- après la méiose : présence de 4 ¢ haploïdes n’ayant qu’un exemplaire de chaque paire de
chromosomes, donc un seul allèle par gène
C. Localisation de la méiose dans le cycle de développement
- cycles de développement : alternance d’une phase haploïde engendrée par la méiose, et d’une phase
diploïde rétablie par fécondation
- Sordaria : méiose immédiatement après fécondation phase haploïde dominante espèce haploïde
- Homme : méiose finie chez l’adulte (parois des tubes séminifères chez l’homme à partir de la puberté,
ovaire chez la femme pendant la vie embryonnaire puis de la puberté à la ménopause s’il y a fécondation),
dans les ¢ de la lignée germinale uniquement phase diploïde dominante espèce diploïde
II - La fécondation permet la formation d’un œuf zygote diploïde
- fécondation : mise en commun des x chromosomes de chaque gamète rétablissement de la diploïdie
- apport de chaque parent de la moitié de son stock chromosomique, donc de la moitié de ses allèles
∟ la fécondation permet donc un mélange aléatoire des allèles des parents
- Homme : pénétration du spermatozoïde dans l’ovaire au stade ovocyte (méiose non finie), fusion des 2
gamètes (caryogamie)
III - Anomalies du nombre de chromosomes
A. Les aberrations chromosomiques
1) Les trisomies (2n+1)
a) La trisomie 21 ou syndrome de Down
- 3 chromosomes 21 caryotype à 47 chromosomes
- phénotype : morphologie faciale différente, nuque courte, handicap mental (amélioration possible par
une éducation adaptée)
- concerne des chromosomes autosomes (non sexuels), comme d’autres trisomies, mais seules les
trisomies 21, 18 et 13 sont viables
b) Le syndrome de Klinefelter
- trisomie qui affecte la paire de chromosomes sexuels XXY
- phénotype : caractères sexuels peu développés, morphologie ambiguë, stérilité
2) Les monosomies (2n-1)
caryotype à 45 chromosomes
- monosomie du chromosome sexuel : X0 = syndrome de Turner
- phénotype : caractères sexuels primaires de la femme, stérilité, pas ou peu de caractères secondaires

B. Origine des aneuploïdies
- accident survenu chez l’un des 2 parents lors de la méiose pendant la gamétogenèse formation de
gamètes anormaux, ayant n+1 ou n-1 chromosomes anomalies du nombre de chromosomes
- accident lorsque 2 chromosomes homologues ou 2 chromatides d’un même chromosome ne se
séparent pas en anaphase I ou II
- fécondation des gamètes anormaux par des gamètes normaux ¢ œufs à 2n+1 ou 2n-1 chromosomes
Le cycle de développement de toutes les espèces à reproduction sexuée est marqué par l’alternance d’une
phase haploïde engendrée par la méiose et d’une phase diploïde rétablie par la fécondation. La durée
relative de ces 2 phases est différente selon les espèces. C’est l’alternance méiose / fécondation qui assure
la stabilité du nombre de chromosomes propre à chaque espèce. Cependant la non-séparation (ou non-
disjonction) au cours de la méiose lors de la formation des gamètes peut conduire chez les espèces
diploïdes à des aberrations de leur nombre.
REPRODUCTION SEXUEE ET VARIABILITE GENETIQUE DES INDIVIDUS
Au sein de l’espèce humaine, le polymorphisme génique concerne environ 10.000 gènes (30% des 30.000
gènes). Les autres gènes possèdent pour la plupart un polyallèlisme, c’est-à-dire un nombre plus ou moins
important d’allèles, sans que chacun soit représenté avec une fréquence significative (supérieure à 1%).
Ainsi chaque individu est hétérozygote pour de nombreux gènes. L’homozygotie complète n’est pas
naturelle mais peut être obtenue artificiellement pour certaines espèces.
I - Méiose et brassage du matériel chromosomique (exemple : Sordaria)
- cas des haploïdes : phénotype à l’image du génotype, individu forcément de race pure
- 2 groupes d’asques obtenus par méiose :
∟ asques de type 4 – 4 : allèles parentaux séparés en méiose I, pas de crossing-over, ségrégation
des allèles parentaux dès la méiose I, 2 types d’asques pré-disjoints équiprobables (disposition des
chromosomes homologues en métaphase I aléatoire)
∟ asques de type 2 – 4 – 2 ou 2 – 2 – 2 – 2 : crossing-over en prophase I au niveau des chiasmas,
ségrégation des allèles en méiose II, chaque noyau issu de la méiose I a les 2 allèles du gène, 4 types
d’asques post-disjoints équiprobables (disposition des chromosomes en métaphase II aléatoire)
- probabilité d’avoir un crossing-over dépendante de la distance entre le centromère et le gène : plus la
distance est grande, plus la probabilité d’avoir un crossing-over est grande
- toujours plus de spores contenant une chromatide non recombinée que recombinée
méiose :
- ségrégation des allèles parentaux en méiose I ou II
- brassage du matériel chromosomique
∟ par le positionnement aléatoire des chromosomes et des chromatides en métaphase
∟ par crossing-over (échanges entre segments de chromatides homologues en prophase I)
II - Brassage allélique par la méiose et par la fécondation chez les espèces diploïdes
A. Cas de loci indépendants (portés par des chromosomes différents)
- détermination de l’indépendance des gènes grâce à un croisement test avec un double bi-récessif
détermination du rôle joué par la méiose : les pourcentages des phénotypes des descendants reflètent
 6
7
6
7
1
/
7
100%