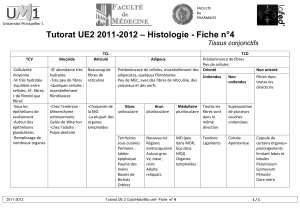
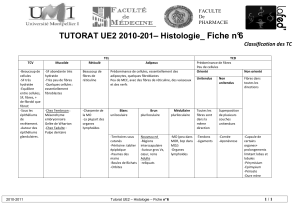
III.3.2. Organe tendineux de golgi.

UE 5 – Physiologie – Chapitre 2 17.09.2012
1
LA SOMESTHESIE
- La somesthésie signifie sensibilité du corps.
- Ces sensibilités sont transportés des récepteurs jusqu’au cortex par deux types de voies :
o La voie lemniscale pour :
o La sensibilité tactile épicritique.
o La sensibilité proprioceptive (sens de position des segments de membres dans l’espace et les uns par
rapport aux autres) => les troubles se révèlent d’abord par des troubles moteurs.
o La voie extra-lemniscale ou faisceau spino-thalamique pour :
o La sensibilité thermique.
o La nociception.
o La sensibilité protopathique.
I. Rappels anatomiques.
- Le cerveau comprend 4 lobes :
o Lobe frontal.
o Lobe pariétal.
o Lobe temporal.
o Lobe occipital.
- Deux scissures :
o Scissures verticale (ou de Rolando) qui délimite le lobe frontal et le lobe pariétal.
o Scissure horizontale (ou de Sylvius) qui délimite le lobe temporal du lobe pariétal et lobe frontal.
- A l’intérieur de chaque lobe on retrouve des circonvolutions (ou gyrus) :
o Gyrus pariétal ascendant (où aboutissent les informations de la somesthésie).
o Gyrus frontal ascendant (où aboutit la voie pyramidale).
- Substance grise :
o Cortex.
o Noyaux gris : thalamus, noyau caudé, noyau lenticulaire.
II. Les grandes voies.
II.1. Voie lemniscale.
- Neurones en T avec corps cellulaires dans le ganglion de la racine
postérieur de la moelle épinière.
- Ce neurone passe par les cordons postérieurs dans faire de relais et
fait un relais dans le noyau Gracilis.
- Le second neurone décusse et monte dans le lemniscus médian puis
fait un relais dans le noyau thalamique ventro-postéro-latéral (relais
de toutes les informations somesthésiques).
- Le 3ème neurone se projette dans le cortex pariétal ascendant en
respectant la somatotopie. (modèle de l’homonculus de Penfield).
- Du plus médian au plus latéral :
o Membre inférieur.
o Tronc.
o Membre supérieur.
o Main. +++
o Face. +++
o Phonation.
- Les informations somesthésiques qui viennent de la face :
o Viennent des nerfs trijumeaux (V).
o Rentre dans le pont.
o Relais dans le noyau du V.
o Relais dans la partie médiane du noyau ventro-postéro-latéral du thalamus.
o Projette sur le cortex pariétal ascendant.

UE 5 – Physiologie – Chapitre 2 17.09.2012
2
II.2. Voie extralemniscale.
- Relais dans la corne grise postérieure. Puis décussation dans la ME au niveau du cordon latéral.
- Puis ça monte dans le faisceau spinothalamique pour se terminer dans le VPL du thalamus, puis le 3ème neurone projette sur
le cortex.
- Faisceau spinothalamique est subdivisé en deux types de faisceaux :
o Voie néo-spinothalamique : assure le transport des informations conscientes (somatotopie).
o Voie paléo-spinothalamique (ou réticulo-spino thalamique) : perte de somatotopie.
o Permet de mettre en alerte le système nerveux. Exemple : pincement quand on dort réveil.
o Met en jeu des mécanismes de contrôle de
l’information douloureuse.
- Stimulation thermique de la face (voie spinothalamique) :
o Information passent par le nerf trijumeau V.
o Passe dans le ganglion trijuminal de Gasser où le corps du
neurone en T.
o Relais dans la partie inférieur du noyau du V dans le tronc
cérébral.
o Décussation.
o Remonte dans le noyau thalamique ventro-postéro-latéral.
o Projette dans la zone de la face du cortex.
II.2.1.Faisceau néo-spinothalamique.
- Relais dans la corne antérieure de substance grise de la moelle épinière.
- Relais dans noyau postéro-ventro-latéral du thalamus.
- Pour le N. trijumeau V :
o Les fibres nociceptives rentrent au niveau de la
protubérance.
o Descendent pour faire relais dans la partie descendante du
noyau du V.
o Relais dans le noyau trijuminal.
o Remontent.
II.2.2. Faisceau paléo-thalamatique.
- 1er relais : corne antérieure de la substance grise de la moelle
épinière.
- Décussation.
- 2ème relais : noyau intralaminaires du thalamus.

UE 5 – Physiologie – Chapitre 2 17.09.2012
3
- Mais envois des collatérales pour faire des projections :
o Au niveau de la réticulée.
o Au niveau du bulbe.
o Au niveau de l’amygdale (structure dans le lobe temporal, impliqué dans la physiologie des émotions).
Des informations douloureuses montent et se projettent sur l’amygdale pour générer des émotions, sans pour
autant se projeter sur le cortex sensitif (information douloureuse peut générer des émotions sans qu’on soit
conscient de la douleur).
o Au niveau de l’insula.
o Au niveau du cortex cingulaire.
o Au niveau de l’hypothalamus : stimulation de l’axe corticotrope. Douleur à l’origine d’une libération de cortisone et
de noradrénaline.
- Ces projections sont impliquées dans les signes neuro-végétatifs qui accompagnent une stimulation douloureuse.
Application expérimentale : stimulation douloureuse sur l’animal à l’origine d’une tachycardie, d’une hypertension, d’une
sudation, mydriase.
II.3.3.Douleur viscérale.
- Trajet particulier.
- Passe dans les ganglions postérieur.
- Relais dans les fibres de la partie très médiane de la substance grise.
- Empreinte les cordons postérieurs.
- Puis croise la ligne médiane au niveau du bulbe.
- Rejoint les thalamus.
- Projection surtout dans l’insula.
- Processus d’inhibition latérale : permet le passage d’une information qui reste très focalisée dans l’espace.
- Les dermatomes radiculaires +++ : voir cours Havet
o Th5 : mamelon
o Th10 : ombilic
- Pour faire une anesthésie il faut couper la racine et celui au-dessus et en dessous
II.4. Résumé.
- Plusieurs composantes dans la douleur :
o Composante perceptive : renseigne sur la localisation de la douleur, intensité, localité fonction du néo-
spinothalamique sur le cortex sensoriel primaire et secondaire (Cortex sensoriel primaire : circonvolution frontale
ascendante).
o Composante affective/émotionnelle et neuro-végétative fonction du système réticulo-thalamique/paléo-spino-
thalamique qui projette sur :
o Le cortex cingulaire antérieur.
o L’insula.
o L’amygdale temporale : structure capitale dans la physiologie des émotions.

UE 5 – Physiologie – Chapitre 2 17.09.2012
4
o L’hypothalamus : responsable de la composante neuro-végétative de la douleur. En cas de
douleur (=situation de stress) il y a une mydriase, hypersudation, tachycardie, etc. qui témoignent de la
mise en jeu du système orthosympathique.
III. Les récepteurs.
III.1. Généralités sur un récepteur.
- La fonction d’un récepteur est de coder l’information c'est-à-dire de transformer différents stimulus naturels en un message
nerveux (constitué par une succession de potentiels d’action).
- Codage de l’intensité : plus la stimulation est intense, plus il élève la fréquence des PA.
- On distingue deux grands types de récepteurs :
o Récepteurs à adaptation lente : toujours actifs durant toute la durée de stimulation. Ils ont une sensibilité
statique : ils codent un état qui se maintient dans le temps.
o Récepteur à adaptation rapide : plus on avance dans le temps d’une stimulation continue plus les potentiels
d’action sont rare. Ils ont une sensibilité dynamique : ils codent des changements d’état.
- Ces deux types de récepteurs qui sont sensible au même type de stimulus ne vont pas envoyer le même type d’information
au cerveau mais sont nécessaire pour coder de manière complète.
Revoir la dépolarisation des cellules (cours de P1)
III.1. Mécanorécepteurs.
- Il en existe 5 types.
- Il y a deux grands types de mécanorécepteurs selon leur fonction :
o Récepteurs à adaptation rapide (sensibilité dynamique essentiellement, sensibles aux variations d’état) qui
comprennent :
o Corpuscules de Pacini :
o Grande sensibilité dynamique (variations de pression).
o Localisation en profondeur du derme.
o Récepteurs de Meissner :
o Grande sensibilité dynamique.
o Localisation superficielle à la limite du derme et l’épiderme.
o Récepteurs liés aux follicules pileux.
o Récepteurs à adaptation lente (sensibilité statique essentiellement, code un état) :
o Disque de Merkel :
Grande sensibilité statique.
Localisation superficielle à la limite du derme et l’épiderme.
o Terminaisons de Ruffini :
Grande sensibilité statique.
Localisation à la profondeur du derme.

UE 5 – Physiologie – Chapitre 2 17.09.2012
5
6. Application clinique
- Compas de Webber : compas à deux pointes qui peuvent être écartés.
- Il permet de tester la sensibilité discriminative : sensibilité qui permet de distinguer deux points.
- Cette sensibilité discriminative est variable d’un endroit à l’autre de la peau, elle est fonction du nombre de récepteurs
par surface.
- Exemple :
o Langue : lieu où il y a le plus de récepteurs tactiles.
o Dos de l’épaule : pour une même surface il y a beaucoup moins de récepteurs.
III.2. Thermorécepteurs.
- Ce sont des récepteurs sensibles au chaud et au froid.
- Ce sont des fibres nerveuses à terminaison libre.
- Récepteurs au chaud : actifs entre 35 et 45°C.
- Récepteurs au froid : actifs entre 10 et 30°C.
- Courbe en cloche (phase ascendante et phase descendante) de ces récepteurs dans un diagramme fréquence de PA en
fonction de la température.
III.3. Les 3 récepteurs de la proprioception.
III.3.1. Fuseau neuromusculaire.
- Sensible aux variations de longueur du muscle (situé en parallèle des fibres musculaires) => excité quand le muscle
s’allonge.
- On trouve des fibres musculaires intra-fusoriales :
o Des fibres à sac nucléaires : noyaux regroupés.
o Des fibres à chaines nucléaires : noyaux à la chaine.
- Elles sont innervées par des fibres afférentes. => double sensibilité :
o Fibres IA ou annulo-spiralées: myélinisées de gros calibre (16 à 20micron) avec corps cellulaire dans la racine
postérieure de la ME (cellules en T) => sensibilité dynamique (aux variations de longueur).
o Fibres secondaires (8 à 12micron) : terminaisons en râteau, elles s’articulent sur les parties proximales et distales
des fibres (sensibilité statique). Corps cellulaires dans la racine postérieure de la ME (cellules en T).
- C’est un récepteur situé en parallèle par rapport aux fibres musculaires striées squelettiques.
 6
7
8
9
6
7
8
9
1
/
9
100%