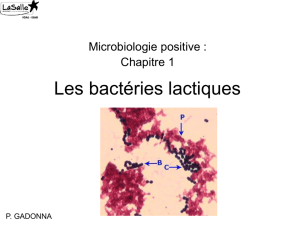
Inhibition des bactéries indésirables par l`activité antimicrobienne

République Algérienne Démocratique et Populaire
Ministère de l'enseignement supérieur et de la recherche scientifique
Université d'Oran-Es-Sénia
Faculté des Sciences
Département de Biologie
Mémoire de Magister
Option: Microbiologie fondamentale et appliquée
Présenté par
Melle : HANNACHI Souad Sabrina
Thème
Inhibition des bactéries indésirables par l’activité
antimicrobienne des espèces de Leuconostoc
isolées du lait cru de chèvre
Devant les membres du jury :
Mr K. El. Aboudi Professeur, Université d'Oran Président
Mr J.E.Henni
Professeur, Université d'Oran Examinateur
Mr T. Sahraoui Maitre de conférences, Université d'Oran Examinateur
Mr M. KIHAL Professeur, Université d'Oran Rapporteur
Melle Z. Benmechernene Docteur, Université d'Oran Co Rapporteur
2007-2008

Remerciements
Ce travail rentre dans le cadre d’un projet CNEPRU ; il a été réalisé a l’université
d’Oran, Faculté des sciences au Laboratoire de Microbiologie appliquée et fondamentale
sous la direction du Pr Kihal Mabrouk. Je voudrais lui exprimer ma profonde gratitude de
m’avoir dirigé tout au long de ce travail avec beaucoup de patience, grâce à ces conseils, ce
travail à été achevé.
J’exprime toute ma gratitude au Dr Benmechernene , co-directeur du mémoire, pour
tout son aide.
Toute ma reconnaissance va au Pr Henni et Dr Sahraoui d’avoir accepté de faire
partie du jury de mémoire en qualité d’examinateur, et au Pr El-Aboudi d’avoir accepté
d’être le président du jury, ainsi que pour l'attention et l'intérêt qu’ils ont portés à ce travail.
La rédaction de ce mémoire me donne l’occasion de remercier ainsi toutes les
personnes qui ont participé et contribué au bon déroulement de ce modeste travail, toutes les
personnes qui de loin ou de prés m’ont aidée dans la réalisation de ce travail ; je cite
quelques noms : Dr Bensalah. F, Mr Yehlalli.Y (de l’université claude bernard France), Mr
AEK et Mr Ahmed (du laboratoire pédagogique de microbiologie Université Es Senia),Mr Ait
Abdeslam N., Mme Hamini..N, Karkachi S et Mme Benhammouche N…
Je remercie, plus particulièrement toute l’équipe de notre laboratoire de
microbiologie appliquée et fondamentale et du laboratoire de phytopathologie
Je remercie chaleureusement ma promotion de Magister ainsi toutes personne qui m’a
soutenue.
Je témoigne toute ma reconnaissance à mon fiancé pour sa disponibilité quotidienne,
ses conseils et son encouragement, toujours là dans les bons et les mauvais moments.
« Merci Maman, merci Papa, je remercie les membres de ma famille pour leur amour »

Dédicace
Je tenais en mon âme à dédire ce fruit de toute une carrière
en signe de respect et d’amour et de reconnaissance aux êtres
les plus chers à mon cur ; Mes parent que dieux les garde pour moi
Une dédicace spéciale à l’être qui m’a donné beaucoup,
sans hésitation à mon cher fiancé
Mr Kamraoui A.
Je dédie ainsi ce travail à ma très chère
sur ;Lemaane ainsi à mes
fréres « Zakkaria, Mehdi et Ayoub »
.

Sommaire
Remerciement
Dédicace
Résumé
Liste des figures
Liste des tableaux
Abréviation
Introduction générale
Introduction 1
Synthèse bibliographique
I-Généralité sur le lait de chèvre 2
1. Définition 2
2. Composition du lait de chèvre 2
II. Généralité sur Leuconostoc 3
1. Définition des Leuconostoc 3
2. Habitat des leuconostocs 4
2.1. Présence dans divers environnement 4
2.2. Présence dans les produits fermentés 5
2.3. Présence en produits laitiers 5
3. Rôle clinique 6
3.1. Infection et épidémiologie 6
3.2. Résistance aux antibiotiques 6
3.3. Les activités métaboliques bénéfiques sur la santé 7
4. Caractérisation et taxonomie 7
4.1. Identification, caractérisation et biodiversité 7
4.2. Taxonomie récente 8
4.3. Comparaison des méthodes d’identification 9
5. Croissance du Leuconostoc 10
5.1. Culture et conservation des cultures 10
5. 1.1. Isolement et milieux de cultures 10

5. 1.2. Conservation des cultures 12
6. Resistance des Leuconostoc au stress 13
7. Métabolisme du Leuconostoc 15
7.1. Utilisation d'hydrate de carbone 15
7.1.1. Transport du sucre 16
7.1.2. Catabolisme du sucre 16
7.2. Métabolisme d'acides organiques 18
7.3. Métabolisme des composés azotés 21
7.3.1. L’exigence des acides aminés 21
7. 3.2. Le système protéolytique 22
7. 4. Métabolisme du Leuconostoc en présence de l'oxygène 23
7. 5. Métabolisme lipidique 24
8. La génétique des Leuconostoc 24
9. Intérêt biotechnologique des Leuconostoc 26
9. 1. Rôles dans la technologie agroalimentaire 26
9. 1.1.Les formations des ouvertures 27
9.1.2. Production d'arôme 28
9. 2. Rôles en nourritures fonctionnelles 29
9. 2. 1. L’effet probiotique du Leuconostoc 29
9.2. 2 Production des polysaccharides et l’effet prébiotiques 29
9. 2. 3 Production de mannitol 30
9. 2.4. Hydrolyse des -galactosides 31
9. 2. 5. Production des vitamines 31
10. Leuconostoc et l'interaction microbienne 31
10.1. Réactions de Co-agrégation 32
10. 2. Croissance du Leuconostoc en cultures mixtes 32
10. 3. La production d’acide organique 33
10.4. Les bactériophages du Leuconostoc 34
10. 5. Les bactériocines produites par Leuconostoc 34
III-Généralité sur Staphylococcus aureus et Bacillus cereus 36
1. Staphylococcus aureus 36
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
1
/
108
100%