CHAPITRE 2 : Stabilité et variabilité du génome d

CHAPITRE 2 : Stabilité et variabilité du génome d’une espèce à reproduction
sexuée.
L’évolution du monde vivant rime avec innovation génétique, donc variabilité du
génome. Mais la stabilité du génome d’une espèce est obligatoire pour que celle-ci
se maintienne dans le monde vivant. Quelles sont les mécanismes qui assurent la
stabilité des génomes ? Comment variabilité et stabilité sont elles géré au sein d’une
espèce ?
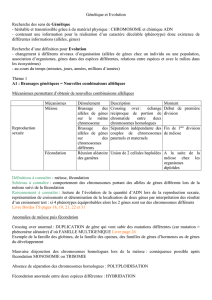
I Cycle des vie ou cycle biologique d’espèces à reproduction sexuée
1) Mécanisme intrinsèques à ces cycles
Il y a 2 mécanismes essentiels à la reproduction sexuée :
-La méiose : Mécanisme permettant la fabrication de gamètes haploïdes (nk) à partir
de cellules souches diploïdes.
-La fécondation : Fusion des gamètes mâles et femelles aboutissant à la formation
d’une cellule œuf diploïde.
Ces 2 mécanismes s’enchainent de génération en génération chez toutes espèces à
reproduction sexuée. Il y a donc dans chaque cycle une phase haploïde et une
phase diploïde.
2) Diversité des cycles biologiques.
-Le cycle humain (cycle de tout mammifère) : phase diploïde dominante
-Cycle d’un champignon ascomycète (cellules reproductrices dans des structures
appelés asques : Ex Sordaria Macrospora)
L’important est de bien comprendre qu’un cycle ne représente pas un individu
mais une espèce. Comme ce cycle se répète, cela illustre la stabilité nécessaire
du génome de l’espèce pour que ce cycle se déroule à nouveau sans
défaillance.
II Analyse du mécanisme de la méiose
1) Approche expérimentale
2) Mécanisme détaillé de la méiose
-Avant toute division il y a une réplication de l’ADN (invisible au microscope)
-Ensuite, division I de méiose :
-Prophase I : Les chromosomes se condensent, l’enveloppe nucléaire disparait.
Particularité de cette prophase : les chromosomes homologues bichromatidiens sont

appariés, ils sont « accolés » leurs chromatides se croisent, cela forme des figures
particulières appelés chiasmas
-Métaphase I : les chiasmas disparaissent, les chromosomes migrent sur le plan
équatorial de la cellule. Ils restent appariés, les centromères sont dirigés vers les
pôles opposés.
-Anaphase I : Grace aux fuseaux de divisions chaque chromosome homologue
migrent vers un pôle opposé de la cellule, on parle de disjonction des chromosomes
homologues.
-Fin anaphase I Télophase I : On obtient 2 lots de n chromosomes bichromatidiens,
en général ils ne vont pas se décondenser complètement car la division II commence
rapidement. La télophase I consiste surtout à séparer la cellule initiale en 2 cellules.
D’une cellule diploïde à 2n k bichromatidiens on obtient 2 cellules haploïdes à n k
bichromatidiens. Cette division permettant de passer de 2n à n k s’appelle division
réductionnelle.
Division II :
-Prophase II : Télescopée avec la télophase I, les chromosomes sont déjà
décondensés.
-Métaphase II : Les chromosomes migrent sur le plan équatorial.
-Anaphase II : Grâce au fuseau il y a séparation des chromatides formant 2 lots de
chromosomes monochromatidiens par cellules.
-Télophase II : Chaque lot se décondense, reformation d’une enveloppe nucléaire,
séparation des cytoplasmes.
De 1 cellule à n chromosomes on passe à 4 cellules à n chromosomes
monochromatidiens. On parle de division équationnelle.
III Reproduction sexuée et stabilité du génome.
1) Méiose et fécondation : 2 rôles complémentaires.
La Méiose est nécessaire dans le cycle biologique, elle permet d’obtenir les gamètes
haploïdes nécessaires à la reproduction. La fécondation a pour rôle de restaurer la
diploïdie par la fusion de gamètes, en particulier la fusion de leur matériel génétique.
Cette dernière s’appelle la caryogamie (fusion des noyaux).Ensemble, méiose et
fécondation permettent une reproduction sexuée qui garantie la stabilité de la ploïdie.
2) Importance de la ploïdie.

Il y a des dysfonctionnements qui peuvent illustrer a quel point la stabilité de la
ploïdie sont importante :
-Trisomie 21: C’est un dysfonctionnement en méiose qui explique l’existence de
gamètes anormaux à 2 chromosomes 21 au lieu d’un seul. Ceci n’arrive que très
rarement. Plus souvent chez la mère que chez le père.
-Un dispositif de la fécondation : Il faut garantir qu’un seul spermatozoïde féconde
l’ovocyte afin que la cellule œuf soit à 2 n chromosomes et pas plus. Or, le
déroulement de la fécondation permet d’éviter la polyspermie (fécondation par
plusieurs spermatozoïdes)
IV Méiose, fécondation et variabilité du génome.
1) La notion essentielle de brassage génétique en Méiose.
Les combinaisons d’allèles des gènes que nous possédons sont normalement les
combinaisons qui étaient présentent dans la cellule œuf. Mais lorsque se déroule la
gamétogénèse, la méiose implique une possibilité de recombinaison des allèles sur
les chromosomes à l’intérieur des nos gamètes. Ce mécanisme s’appelle le
brassage génétique, il ne fonctionne que pour les gènes pour lesquels l’individu est
hétérozygote.
Homozygote pour un gène: avoir le même allèle pour un gène.
Hétérozygote pour un gène : l’allèle paternel est différent de l’allèle maternel pour un
gène donné.
2) Le brassage intra chromosomique.
Le brassage intrachromosomique permet de recombiner des allèles de gènes portés
par une même paire de chromosomes homologues. En génétique, 2 gènes qui sont
sur le même chromosome sont appelés gènes liés. Soient 2 gènes liés A et B, soit
l’individu hétérozygote pour les deux gènes, son génotype est (A1B1//A2B2).

Les combinaisons d’allèles dans les 4 gamètes obtenues sont les mêmes
combinaisons que les combinaisons parentales initiales. Il y a eu aucune
recombinaison, aucun brassage génétique. On dit qu’on obtient 4 gamètes
parentaux. Cela dit il pourrait très bien avoir une méiose avec crossing-over.
Grâce à l’étude du champignon Sordaria, nous savons qu’il peut y avoir lors de la
prophase I un échange de segments de chromatides entre 2 chromosomes
homologues apparié. Ce mécanisme s’appelle le crossing-over, et dans notre cas il
peut permettre une recombinaison des gènes des allèles A et B.
Méiose avec crossing-over

Le mécanisme de brassage ainsi illustré est appelé brassage intrachromosomique. Il
est basé sur le phénomène de crossing-over qui intervient en prophase I.
Chez Sordaria les asques 2-2-2-2 et 2-4-2 obtenues par crossing-over sont plus
rares que les asques 4-4 obtenues sans crossing-over. Le crossing-over étant un
phénomène relativement rare, les méioses avec C.O sont plus rares que les
méioses sans C.O donc les gamètes recombinés sont toujours plus rare que les
gamètes parentaux dans le brassage intrachromosomique. Plus deux gènes sont
liés, sont proches sur le même chromosome, moins il y a de probabilité de séparer
les allèles par C.O.
3) Le brassage interchromosomique.
Ce deuxième mécanisme de brassage génétique permet de recombiner les allèles
de gènes indépendants : sur 2 chromosomes d’une paire différentes. Le mécanisme
est très simple, au cours de la méiose en Anaphase I le chromosome maternel d’une
paire migre indépendamment avec le chromosome maternel ou paternel d’une autre
paire. On dit qu’il y a disjonction indépendante des chromosomes homologues de
paires différentes. Se qui a pour conséquence une ségrégation indépendante des
allèles paternels et maternels de gènes indépendants.
 6
7
8
6
7
8
1
/
8
100%