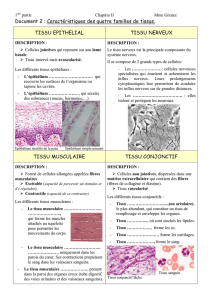
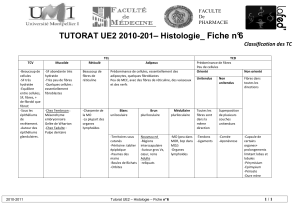
I. Le système tégumentaire

La peau est un organe composé de différents tissus. L’épiderme est un épithélium de
revêtement et le derme un tissu conjonctif.
Epiderme
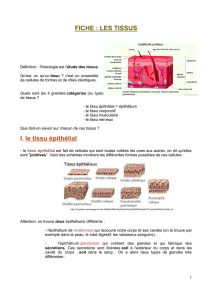
Caractéristiques des épithéliums de revêtement
Les épithéliums représentent un groupe de tissus qui revêtent toute la surface du corps, les
cavités et les tubes. Ils réalisent des interfaces entre les différents compartiments de
l'organisme.
Les épithéliums sont impliqués dans de nombreuses activités comme la diffusion sélective,
l'absorption, la sécrétion, la protection mécanique et le cloisonnement.
Certains épithéliums remplissent plusieurs rôles, ainsi l'épithélium intestinal est spécialisé
dans l'absorption des produits de digestion mais se protège du contenu intestinal nocif en
sécrétant un mucus à sa surface.
Les surfaces épithéliales sont constituées d'une ou plusieurs couches de cellules séparées par
un matériel intercellulaire réduit. Les cellules épithéliales sont étroitement jointes grâce à des
jonctions cellulaires qui permettent le passage d'informations et de métabolites.
Tous les épithéliums reposent sur une membrane basale d'épaisseur variable. Celle-ci sépare
les épithéliums du tissu conjonctif sous-jacent et n’est jamais traversée par des vaisseaux
sanguins; les épithéliums dépendent donc des tissus sous-jacents à partir desquels diffusent
l'oxygène et les métabolites.
1
I. Le système tégumentaire

On distingue deux grands groupes d'épithéliums: les épithéliums de revêtement et les
épithéliums glandulaires.
Les cellules d'un épithélium de revêtement sont étroitement associées grâce à des jonctions
intermembranaires spécialisées: les desmosomes. Ceux-ci s'accompagnent de tonofilaments
cytoplasmiques qui renforcent les adhérences entre cellules adjacentes.
Une membrane basale faite de mucopolysaccharides sépare le tissu épithélial du tissu
conjonctif voisin.
Le tissu épithélial de revêtement limite une cavité ou recouvre un organe et repose toujours
sur un tissu conjonctif appelé chorion.
Le tissu épithélial n'est pas vascularisé mais est parfois richement innervé.
Sa capacité de régénération est grande; les cellules de ces tissus devant souvent être
renouvelées.
Classification des épithéliums de revêtement
On classe ces épithéliums selon trois caractères morphologiques:
a) le nombre de couches cellulaires:
- un épithélium comportant une seule couche de cellules est appelé unistratifié,
- un épithélium comportant plusieurs couches cellulaires est appelé pluristratifié.
b) la forme des cellules observées sur des coupes perpendiculaires à la surface épithéliale:
pour les épithéliums pluristratifiés, la classification est déterminée par la forme de la couche
la plus externe.
c) la présence de spécialisations de structure tels les cils ou la kératine: ainsi, l'épithélium de
surface de la peau est classé comme "épithélium pavimenteux pluristratifié kératinisé".
Epithéliums unistratifiés:
Ces épithéliums simples sont formés d'une seule couche de cellules. On les trouve presque
toujours au niveau d'interfaces impliquées dans la diffusion sélective, l'absorption ou la
sécrétion. Ils constituent une faible protection contre l'abrasion et on ne les rencontre
presque jamais sur les surfaces soumises à une forte agression.
2

Les cellules de l'épithélium unistratifié peuvent être cubiques, aplaties ou pavimenteuses,
cylindriques, selon leur fonction.
Les épithéliums pavimenteux simples constituent une faible barrière à la diffusion et sont
donc localisés au niveau des alvéoles pulmonaires, de la bordure des vaisseaux sanguins
(endothélium) et des cavités du corps (mésothélium).
Les cellules de cet épithélium sont aplaties, de forme irrégulière et constituent une surface
continue.
Les épithéliums pavimenteux unistratifiés bordent souvent des surfaces spécialisées dans
les transports passifs de gaz ou de liquides. Ils forment aussi la fine bordure des cavités
pleurale, péricardique et péritonéale et permettent le passage des fluides d'origine
tissulaire vers l'intérieur ou vers l'extérieur des ces cavités.
Les épithéliums cubiques simples représentent une forme intermédiaire entre les
épithéliums pavimenteux simples et les épithéliums cylindriques simples. Sur des coupes
perpendiculaires à la membrane basale, les cellules apparaissent carrées.
Le noyau est habituellement arrondi et localisé au centre de la cellule.
Les épithéliums cubiques simples bordent habituellement des petits canaux et des tubules
qui ont des fonctions d'excrétion, de sécrétion ou d'absorption comme les petits canaux
collecteurs du rein, des glandes salivaires et du pancréas.
Les épithéliums cylindriques simples sont formés de cellules plus hautes que larges
apparaissant cylindriques sur des coupes perpendiculaires à la membrane basale.
Les noyaux sont allongés et se localisent souvent à la base des cellules, c'est à dire à leur
pôle basal. Le pôle apical est parfois modifié et peut présenter des cils, des stéréocils, des
microvillosités ou une substance sécrétée par une cellule à différenciation glandulaire.
L'épithélium cylindrique simple se trouve essentiellement au niveau des surfaces à rôle
d'absorption comme l'intestin grêle mais peut également revêtir des muqueuses sécrétrices
comme celles de l'estomac.
L’épithélium cylindrique simple cilié se caractérise par la présence de cils au pôle apical
des cellules.
Ces spécialisations de surface sont facilement visibles en microscopie optique. Parmi les
cellules ciliées, on observe quelques cellules dispersées non ciliées qui ont, habituellement
3

un rôle sécrétoire. Chaque cil consiste en une expansion de la membrane plasmique en
forme de doigt, contenant une spécialisation mobile du cytosquelette. Chaque cellule peut
présenter jusqu'à 300 cils, qui se balancent et créent un courant permettant aux fluides et
aux petites particules de se déplacer à la surface de l'épithélium.
L'épithélium cylindrique simple cilié est visible au niveau de tractus génital féminin.
Il est probable que les cils de la muqueuse tubaire créent un courant favorisant le transport
de l'ovule depuis l'ovaire jusqu'à l'utérus.
L’épithélium cylindrique simple cilié pseudostratifié tire son nom du fait que, sur une
section, l'épithélium peut apparaître, de façon erronée, pluristratifié. Il s'agit d'un
épithélium unistratifié simple puisque toutes les cellules reposent sur une membrane
basale. Les noyaux de ces cellules sont disposés à différents niveaux et donnent ainsi
l'illusion d'une stratification cellulaire.
On ne trouve d'épithélium pseudostratifié que presque exclusivement au niveau des voies
aériennes supérieures chez les mammifères: on l'appelle souvent épithélium respiratoire.
Remarque: La transition entre deux types d'épithéliums peut être brusque, comme entre
l'oesophage et l'estomac ou progressive, comme entre deux épithéliums de revêtement.
Epithéliums pluristratifiés:
Ces épithéliums se caractérisent par la présence de deux ou plusieurs couches cellulaires.
Ils ont une fonction de protection; le degré et la nature de la stratification dépendent des
différentes sortes d'agressions physiques auxquelles la surface est exposée.
On classe les épithéliums pluristratifiés d'après la forme des cellules les plus
superficielles; toutes les cellules basales étant en général de forme cubique.
On distingue les épithéliums pluristratifiés suivants:
a) pavimenteux épidermique
b) pavimenteux épidermoïde
c) cubique
d) cylindrique
e) urinaire
4

Les Epithéliums pluristratifiés pavimenteux sont constitués d'un nombre variable
d'assises cellulaires qui subissent une évolution morphologique et fonctionnelle, depuis
l'assise basale cubique jusqu'aux couches superficielles.
Les cellules basales engendrent, par mitoses successives, des cellules qui vont
progressivement gagner la surface où elles desquament. Pendant leur migration vers la
surface, les cellules subissent tout d'abord une maturation puis une dégénérescence au fur
et à mesure qu'elles s'éloignent du tissu conjonctif nourricier sous-jacent. Cet épithélium
est bien adapté pour résister à l'abrasion; il se trouve au niveau de la cavité buccale, du
pharynx, de l'oesophage, du vagin.
Les différentes couches cellulaires sont les suivantes:
a) couche basale: cellules cubiques de régénération;
b) couche moyenne: cellules polyédriques nucléées;
c) couche superficielle: cellules aplaties nucléées sans kératinisation et qui desquament.
L' Epithélium pluristratifié pavimenteux kératinisé
L'épiderme est une variété d'épithélium pavimenteux pluristratifié adapté aux phénomènes
constants d'abrasion et de dessiccation auxquels la surface du corps est exposée. Pendant
la maturation, les cellules épithéliales subissent le phénomène de kératinisation,
aboutissant à la formation d'une couche épaisse formée d'une protéine: la kératine.
L'épithélium comprend le corps muqueux de Malpighi et une couche cornée.
Dans le corps muqueux de Malpighi, on trouve:
a) une couche de cellules cubiques basales, sans cesse renouvelées. Ces cellules
contiennent un pigment: la mélanine.
b) une couche moyenne de cellules polyédriques nucléées.
c) une couche granuleuse comportant des cellules aplaties anucléées dont le cytoplasme
est encombré de gros grains de kérato-mélanine.
Dans la couche cornée, les cellules ont perdu leur noyau. Cette couche est épaisse au
niveau des doigts et de la plante des pieds.
Les Epithéliums pluristratifiés cubiques sont fins et comprennent deux ou trois couches
de cellules cubiques ou cylindriques basses. Ils revêtent les canaux excréteurs de glandes
5
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
1
/
38
100%