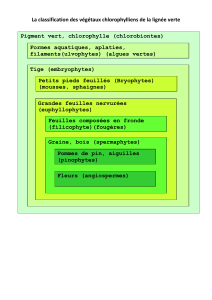
systématique des embryophytes

Université Pierre et Marie Curie (PARIS 6)
Préparation à l’agrégation SVSTU, secteur B
SYSTÉMATIQUE DES EMBRYOPHYTES
Document illustrant plus particulièrement les taxons européens
version 4 octobre 2007
Catherine Reeb, Préparation agrégation SVSTU, UPMC
Jean-Yves Dubuisson, Laboratoire Paléobotanique et Paléoécologie, équipe Paléodiversité, systématique et évolution
des Embryophytes, UPMC

Dessin de couverture: extrait d’un traité botanique tibétain
Pour Garance
Remerciements:merci à Jean et Monique Duperon pour les clés de détermination des familles d’Angiospermes, à
A.M pour la reproduction de couverture, à Michaël Manuel et Eric Queinnec pour la partie I de l’introduction. Egalement
à tous les illustrateurs qui ont autorisé l’utilisation de leurs documents mis en ligne sur Internet: Françoise Gantet, Prof.
I. Foissner, Association Endemia de Nouvelle Calédonie.
1

Table des matières
Introduction 3
Les données essentielles pour l’agrégation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
I Systématique générale des Embryophytes 6
I.1 CaractèresdesEmbryophytes......................................... 6
I.2 La place des Embryophytes au sein de la lignée verte . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7
I.3 LegroupefrèredesEmbryophytes...................................... 7
I.4 Relations au sein des Embryophytes:les points acquis aujourd’hui . . . . . . . . . . . . . . . . . . . . . 8
I.4.1 Marchantiophytes, Bryophytes, Anthocérotophytes à la base des Embryophytes . . . . . . . . . . 8
I.4.2 Les Trachéophytes ou plantes vasculaires, un groupe monophylétique . . . . . . . . . . . . . . . 9
I.4.3 Les Spermatophytes ou plantes à ovules, un groupe monophylétique . . . . . . . . . . . . . . . . 9
I.5 Quelques points encore en discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
I.5.1 Position et relations des trois clades basaux (Bryophytes, Marchantiophytes et Anthocérotophytes) 11
I.5.2 Relations au sein des Spermatophytes:l’hypothèse "Gnepine" . . . . . . . . . . . . . . . . . . . 13
II Les principaux clades d’Embryophytes 15
II.1 MARCHANTIOPHYTA:Les Marchantiophytes ou Hépatiques . . . . . . . . . . . . . . . . . . . . . . 15
II.2 ANTHOCEROTOPHYTA:Les Anthocérotophytes ou Anthocérotes . . . . . . . . . . . . . . . . . . . . 17
II.3 BRYOPHYTA:Les Mousses au sens strict:mousses et sphaignes . . . . . . . . . . . . . . . . . . . . . . 18
II.4 POLYSPORANGIOPHYTA:Les Polysporangiophytes . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
II.5 TRACHEOPHYTA:Les Trachéophytes ou plantes terrestres . . . . . . . . . . . . . . . . . . . . . . . . 22
II.6 EUTRACHEOPHYTA:Les Eutrachéophytes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22
II.7 LYCOPHYTA:LesLycophytes........................................ 24
II.8 EUPHYLLOPHYTA:Les Euphyllophytes, végétaux vasculaires à croissance pseudomonopodiale . . . . . 27
II.9 MONILIFORMOPSES = MONILOPHYTES . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29
II.10 EQUISETOPHYTA:les Equisetophytes ou Sphenophytes ou prêles . . . . . . . . . . . . . . . . . . . . 30
II.11Fougèresleptosporangiées .......................................... 31
II.12 Un groupe paraphylétique:les fougères eusporangiées. . . . . . . . . . . . . . . . . . . . . . . . . . . . 35
II.13LIGNOPHYTA:lesLignophytes....................................... 35
II.14 SPERMATOPHYTA:les Spermatophytes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35
II.14.1 Les Ptéridospermales = fougères à graines, taxon para- ou polyphylétique . . . . . . . . . . . . . 37
II.15CYCADOPHYTA:lesCycadophytes..................................... 37
II.16GINKGOPHYTA:lesGinkgophytes ..................................... 39
II.17 LES CONIFÈRES...des relations en cours de résolution. . . . . . . . . . . . . . . . . . . . . . . . . . . 40
II.17.1 Détails des principaux taxons de Conifères, hors Gnetophytes . . . . . . . . . . . . . . . . . . . 41
II.18GNETOPHYTA:lesGnetophytes....................................... 45
II.19 ANGIOSPERMES:les plantes à fleurs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45
II.19.1L’originedesAngiospermes ..................................... 46
II.19.2 Les systèmes de classification traditionnelles: . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
II.19.3 Commentaires sur les principaux clades d’Angiospermes. . . . . . . . . . . . . . . . . . . . . . 49
II.19.4 Quelques espèces d’Angiospermes communes ou présentant un intérêt particulier (agronomique,
scientifiqueetc). ........................................... 53
III Clés de détermination 55
III.1 Clés de détermination des principaux taxons d’Embryophytes . . . . . . . . . . . . . . . . . . . . . . . . 55
III.2 Clé des familles européennes de Moniliformopses avec genres représentatifs et quelques exotiques . . . . 57
III.3 Clé des familles actuelles de Coniferophytes avec genres représentatifs . . . . . . . . . . . . . . . . . . . 59
III.4Clédesfamillesd’Angiospermes....................................... 60
III.4.1 PlantesDicotylédones ........................................ 60
III.4.2 PlantesMonocotylédones ...................................... 62
2

Introduction
Ce document, notamment son introduction dont nous reprenons in extenso la plupart des paragraphes, a été largement
inspiré du polycopié "Systématique des métazoaires, 2002 " de Michaël Manuel et Eric Queinnec, que nous remercions
vivement ici.
Objectifs de ce document
Ce document a pour objectif de replacer dans une classification phylogénétique des taxons, regroupant tous les "végé-
taux " terrestres, dont certains sont secondairement revenus au milieu aquatique. Nous y incluons les taxons observables
en France, et quelques taxons tropicaux d’intérêt agronomique dont la connaissance pourra être exigée pour le concours
de l’agrégation. Le terme de "végétaux " a été volontairement mis entre guillemets, car si les animaux recouvrent assez
naturellement le taxon des Métazoaires, il n’en est pas de même pour les organismes prétendant au statut de "végétal ".
Selon les caractères choisis pour les définir, les taxons inclus ne sont pas les mêmes. Les Embryophytes recouvrent par
contre un groupe naturel, monophylétique.
La présentation de la classification phylogénétique des Embryophytes s’accorde tout à fait avec l’esprit de réforme du
programme de l’Agrégation, dont un des principaux aspects vise à mieux intégrer les connaissances "naturalistes " au
sein des grandes problématiques évolutives, biologiques, écologiques etc. De ce point de vue, il est clair que la biologie
fonctionnelle et la systématique, loin de s’opposer, s’éclairent mutuellement, et c’est pourquoi les données biologiques
ont été intégrées dans la présentation des taxons, autant qu’il était possible dans la limite de la place disponible.
Principes sous-jacents
L’ambition principale de la systématique contemporaine avait déjà été expriméepar Darwin: les classifications doivent
rendre compte de l’histoire évolutive des organismes. En effet tout autre critère de classification serait arbitraire et artificiel.
Seulement, que savons nous de cette histoire évolutive, puisque nous n’étions pas là? Reconstituer l’histoire à partir de
l’héritage du passé, c’est cela l’objectif de la phylogénie. L’héritage du passé comprend
–la diversité des êtres vivants actuellement observables sur notre planète,
–les archives paléontologiques, c’est à dire les fossiles.
La véritable révolution systématique dans la seconde moitié du XXème siècle, ce n’est pas, comme on pourrait le
penser, l’utilisation des données moléculaires, mais bien l’adoption et les méthodes de la cladistique, progressivement
par l’ensemble des chercheurs en phylogénie, à la suite des travaux de Hennig (à partir des années 60). Auparavant, les
systématiciens imaginaient des scénarios évolutifs sans véritable méthodologie; c’était une activité essentiellement spécu-
lative où prévalait l’opinion personnelle sur l’importance de tel ou tel caractère, ou sur telle modalité supposée d’évolution
des caractères et des taxons. On privilégiait volontiers les hypothèses d’évolution complexifiante, et du coup on classait
souvent les organismes en "groupes primitifs " et en "groupes évolués" comme si l’évolution était synonyme de com-
plexification, et comme si le taxon ne pouvait présenter à la fois des caractères primitifs et en même temps des caractères
évolués. On comprend aujourd’hui que cette manière de penser est totalement inappropriée, car il s’est écoulé exactement
le même temps entre la première cellule (il y a plus de 3,8 milliards d’années) et n’importe lequel des organismes ac-
tuels, quel que soit son "degré de complexité " apparent. Des transformations évolutives se sont produites dans toutes les
lignées, dans toutes les branches de l’arbre du vivant; ces transformations sont, selon les cas, l’apparition, la perte, ou la
transformation d’un caractère.
C’est cette histoire que les systématiciens d’aujourd’hui tentent de reconstituer en appliquant la méthode cladistique.
Insistons sur le fait qu’ils utilisent tout aussi bien les caractères morphologiques (incluant ceux qui sont fournis par les
techniques de microscopie électronique), que des caractères biochimiques ou moléculaires (séquences de gènes, organi-
sation des génomes etc. )
On peut rappeler ici les grands principes de la science moderne des classifications en quelques points (voir l’introduction
de ?pour une présentation détaillée):
Chercher à construire la phylogénie c’est chercher à identifier des groupes monophylétiques (= des
unités de descendance).
Seul le partage des états dérivés de caractères (= les synapomorphies) révèle la monophylie des groupes. Jusqu’à
l’adoption du cladisme, on se basait indistinctement sur les homologies (= caractère hérité d’un ancêtre commun); aujour-
d’hui on distingue au sein de l’homologie, d’une part le partage d’états primitifs (ou ancestraux), qui n’est pas révélateur
Systématique des Embryophytes - version 4 octobre 2007 - C.Reeb et JY. Dubuisson 3/64

de la monophylie, et d’autre part le partage d’états dérivés, qui lui révèle la monophylie (les nouveautés évolutives ac-
quises chez un ancêtre étant héritées par tous ses descendants).
L’analyse cladistique permet de résoudre les conflits entre les caractères, par l’application du principe de parcimonie.
Cette analyse conduit à préférer l’hypothèse phylogénétique la plus parcimonieuse, considérée comme la plus vraisem-
blable, compte tenu de la distribution observée de l’ensemble des caractères (on ne choisit donc plus arbitrairement tel ou
tel caractère pour découper les groupes). En même temps, à partir de l’hypothèse phylogénétique résultant de l’analyse,
il est possible de reconstituer l’histoire de chacun des caractères pris indépendamment. C’est ainsi qu’on peut identifier
les synapomorphies qui soutiennent la monophylie de tel ou tel groupe, et aussi distinguer les caractères qui évoluent de
manière convergente (= homoplasies).
Il n’y a pas de "groupes primitifs " ni de groupes évolués; les qualificatifs "primitif " (= ancestral) / "évolué " (=dérivé)
s’appliquent aux caractères (ou ensembles de caractères) et non aux groupes. Chaque organisme est une mosaïque de ca-
ractères évoluant plus ou moins indépendamment les uns des autres. Seuls les groupes monophylétiques ont une histoire
évolutive propre (les transformations évolutives ayant lieu dans la branche commune -les synapomorphies- concernent
l’ensemble des membres du groupe, et aucun des organismes extérieurs au groupe). C’est pour cette raison qu’on ne re-
tient comme unité de classification, comme taxon (= groupe formellement reconnu dans la classification), que des groupes
dont la monophylie est soutenue par les résultats d’un ou plusieurs travaux de phylogénie.
Il faut insister sur le fait que l’analyse des caractères est explicite et reproductible, ce qui signifie que n’importe qui,
à partir des mêmes données, en appliquant exactement les outils indiqués par les auteurs (dans la partie Matériels et Mé-
thodes de la publication), obtiendra le même arbre. Par ailleurs, à partir de nouvelles données, ou en codant différemment
les caractères, ou en changeant les modalités de l’analyse, on pourra éventuellement contredire une hypothèse phylogéné-
tiquement pré-existante au profit d’une autre, mais toujours à partir d’une démarche explicite (et non pas, en principe, du
fait de convictions personnelles ou d’avis a priori sur l’évolution du groupe).
Ainsi, les classifications proposées dans ce document sont des résultats scientifiques (issus de travaux publiés dans
des revues spécialisées ou des ouvrages synthétisant ces travaux), et l’on doit appliquer le même esprit critique qu’envers
n’importe quel résultat scientifique, comme dans n’importe quelle discipline. Par ailleurs, un résultat scientifique n’est
intéressant que dans la mesure où il apporte du nouveau, et donc on ne doit pas s’alarmer outre mesure du fait que les
classifications changent: c’est même plutôt un signe de vigueur (cela signifie que les systématiciens travaillent!). Il peut
aussi y avoir des contradictions entre les résultats provenant d’auteurs différents et/ou de jeux de données différents: il
y a alors des questions en suspens qui nécessitent d’autres travaux. Une fois établis les groupes monophylétiques, donc
les taxons, on doit les nommer. L’ensemble des règles qui gouvernent l’attribution des noms aux taxons s’appelle la no-
menclature. L’existence de règles est indispensable pour préserver un maximum la stabilité des noms face aux nombreux
travaux des systématiciens et aux changements perpétuels des conceptions sur le découpage des groupes.
Actuellement est toujours en vigueur le code de nomenclature hérité des XVIII ème et XIX ème siècles, qui prévoit
en particulier qu’à chaque taxon soit associée une catégorie linnéenne (les principales catégories sont l’espèce, le genre,
la famille, l’ordre, la classe, l’embranchement ou phylum). Il existe parmi les systématiciens un débat pour savoir si
ce système de nomenclature est adapté aux nouveaux concepts et méthodes issus de la cladistique. Les tenants d’un
nouveau code de nomenclature (le phylocode, consulter http://www.ohio.edu/phylocode) proposent une nouvelle approche
taxonomique.
Il n’est pas encore reconnu, surtout en biologie végétale, et la nomenclature traditionnelle issue de la nomenclature
binomiale est conservée ici. Mais attention, les catégories taxonomiques, familles, ordres n’ont pas d’autres significations
que leur monophylie et le fait qu’ils incluent d’autres groupes monophylétiques. Afin de clarifier cette hiérarchie, des ter-
minaisons appropriées ("phyta " pour les embranchements, "psida " pour les classes, "ales " pour les ordres, "acées " pour
les familles) sont utilisées selon la profondeur du groupe dans la phylogénie. Mais ces groupes n’ont pas de signification
absolue, ni d’équivalence de hiérarchie. Des taxons de même rang ne doivent pas être vus comme des équivalences d’une
réalité biologique. Cette hiérarchie possède avant tout un intérêt didactique et mnémotechnique. C’est une des différences
importantes avec la vision des familles des classifications antérieures.
Guide de lecture
Les chapitres sont construits selon un plan standardisé: une page présentant les caractères du taxon faisant l’objet du
chapitre une hypothèse phylogénétique (dérivée de la confrontation des hypothèses publiées récemment), des remarques
concernant l’arbre (hypothèses alternatives, caractères problématiques, etc. ), puis une section "exemples " illustrant les
taxons terminaux (= les feuilles) de l’arbre. Parfois un ou plusieurs taxons terminaux font l’objet d’un sous-chapitre spé-
cial, dans ce cas ils ne sont pas illustrés dans la partie "exemples " du premier arbre.
Systématique des Embryophytes - version 4 octobre 2007 - C.Reeb et JY. Dubuisson 4/64
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
1
/
65
100%