Page de pre*sentation d`une rone*o

UE11 – Parcours 3– Signalisation
Cellulaire – Cours n° 5
23/03/2016
Pr Vincent Goffin
vincent.goffin@inserm.fr
RT : Jérôme Langlois
RL : Alexandre Meunier
Récepteurs prolactine, hormone de croissance
et récepteurs cytokines
Plan :
1. Aspects moléculaires
A- Présentation des acteurs en présence (ligands, récepteurs)
B- Interaction ligand-récepteur : la première étape
C- Transmission du signal intracellulaire
2. Aspects physiopathologiques
A- Pathologies liées à des anomalies (génétiques) de signalisation
1. Défaut de signalisation
2. Excès de signalisation
B- Approches thérapeutiques
Abréviations :
Hormone de croissance : GH
Récepteur de l’hormone de croissance : GHR
Prolactine : PRL
Récepteur de la prolactine : PRLR
Placental Lactogen : PL
Récepteur de cytokines hématopoïétiques de classe I : RCHCI
Facteur de croissance : GF
Mots du RT et du RL
Faites bien attention aux abréviations employées : elles peuvent être source de confusion. Ce sont
celles utilisées par le professeur pendant son cours ; il faut donc se familiariser avec !

I. Aspects moléculaires
A- Présentation des acteurs en présence (ligands, récepteurs)
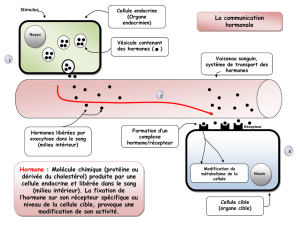
L’hormone de croissance (GH) et la prolactine (PRL) sont des hormones hypophysaires. Ces
deux hormones sont produites dans l’anté-hypophyse. La GH est produite par les cellules
somatotropes et la PRL est produite par les cellules lactotropes.
Ces deux hormones sont sous contrôle d’une fonction régulatrice :
La production de prolactine est inhibée par la dopamine.
La production de GH est inhibée par la somatotropine.
La dopamine et la somatotropine sont produites dans l’hypothalamus.
La GH possède différentes fonctions :
Elle est impliquée dans la croissance et la morphogénèse des os et des tissus mous.
Elle est impliquée dans le métabolisme à travers la lipolyse et la synthèse protéique.
La prolactine est impliquée dans :
La lactation et la reproduction chez l’être humain.
Le comportement et l’immunomodulation chez d’autres espèces.
La balance eau-électrolytes chez les poissons.
Les récepteurs de la GH et de la PRL sont largement exprimés dans un grand nombre de
tissus différents. Ainsi, ces hormones ont de nombreuses fonctions à travers tout l’organisme.
La GH et la prolactine font partie d’une même famille, comprenant également le placental
lactogen (PL). Le PL est une hormone ayant un rôle dans la lactation, comme la prolactine, à la
différence près qu’il s’agit d’une hormone produite par le placenta. Elle est de ce fait exclusive aux
mammifères.
La GH et la PRL sont des hormones anciennes et conservées dans l’évolution. Il n’y avait à
l’origine qu’un seul gène ancestral, avant la divergence avec les poissons.
De même, s’est mis en place, avec le temps, au sein des mammifères, deux types de PL :
Le PL des primates est issu de la GH ;
Le PL des non-primates est issu de la PRL.
Ces deux hormones ont sensiblement la même fonction, mais possèdent un parcours
d’évolution différent. Ce phénomène s’appelle une convergence fonctionnelle.

La PRL et de la GH ont tous les deux une structure 3D qui leur est propre. Elles sont
structurées en faisceaux α de 4 hélices. Il s’agit de la signature structurale de la famille des
cytokines hématopoïétiques.
Les deux hormones sont virtuellement identiques et possèdent une structure similaire ; elles
comprennent 200 acides aminés. Ces hormones peuvent subir des modifications post-
traductionnelles, à savoir phosphorylation, glycosylation, agrégation et protéolyse, lesquelles ont un
impact sur les propriétés biologiques des hormones.
On distingue des cytokines hématopoïétiques à longues chaînes et à courte chaîne :
Longues chaînes : GH, PRL, interférons…
Courtes chaînes : IL-2, IL-4, IL-5…
Il existe 50 membres dans la superfamille des récepteurs de cytokines hématopoïétiques,
répartis en deux classes :
Les récepteurs de classe I (GH/PRL).
Les récepteurs de classe II (interférons).
Les récepteurs de cytokines hématopoïétiques de classe I ont certaines caractéristiques particulières, à
savoir :
Leur partie N-ter est extracellulaire ; leur partie C-ter est intracellulaire.
Le domaine extracellulaire contient 4 Cystéines réparties en une paire de 2 ponts
disulfures séparés. Ces deux paires de cystéines sont primordiales pour maintenir la
structure de la partie extracellulaire du récepteur et ainsi assurer la liaison du récepteur à
l’hormone.
Toujours dans le domaine extracellulaire du récepteur mais plus proche de la membrane,
se trouve un motif WS jouant un rôle dans la structure du domaine extracellulaire et
permettant également l’adressage du récepteur à partir du lieu de synthèse vers la
membrane plasmique. Ces motifs présentent des différences en fonction du ligand.
- Motif du récepteur de la PRL : Trp - Ser - X - Trp - Ser.
- Motif du récepteur de la GH : Tyr - Gly - X - Phe - Ser.
Du côté intracellulaire et proche de la membrane plasmique se situe une région riche en
proline, qui se nomme, la Boîte 1. Cette région est indispensable pour la transduction du
signal.
Toujours du côté intracellulaire, on trouve des motifs qui sont importants pour
l’internalisation du récepteur après exposition à la membrane afin d’être recyclé ou dégradé.
On trouve un motif d’interaction avec le système ubiquitine-protéasome, et un motif
leucine-leucine.
Cystéines
Motifs WS
Boîte 1

La signature structurale des récepteurs de de cytokines hématopoïétiques de classe I est
caractérisée par la présence au niveau extracellulaire de deux sandwich β constitués chacun de 7
brins β.
Il est important de préciser que chaque type de récepteur est codé par un seul gène. En
revanche, un même gène peut donner plusieurs isoformes de ce même récepteur, notamment à
travers l’épissage alternatif. Ainsi, dans le cas de PRLR, on distingue 5 isoformes avec une
pertinence physiologique, qui vont moduler l’action de la protéine. La possibilité de création des
isoformes est permise par l’épissage alternatif des différents exons du gène de la PRLR.
Pour ΔS1, forme où les 2 PDS sont absents, on a une baisse de l’affinité pour la protéine.
Pour les formes S1a, S1b, ou encore intermédiaire, on a une baisse de la transmission du
signal du fait de la troncature de la partie C-terminale.
Enfin, pour la forme soluble, la partie intracellulaire est absente, le récepteur n’est pas
attaché au sein de la membrane plasmique. Il dérive librement dans le lait et vient fixer la
PRL. Ainsi, il augmente la demi-vie de cette dernière, ce qui a fortiori, permet une durée
d’action plus longue.
B- Interaction ligand-récepteur : la première étape.
Il est important de préciser qu’il existe une spécificité de liaison des récepteurs aux hormones :
Le GHR ne fixe que la GH. Il est néanmoins possible de créer un GHR muté liant la PRL,
en agissant sur 6 acides aminés.
Le PRLR fixe la GH, le PL, la PRL. Il est néanmoins possible de créer un PRLR muté sans
affinité pour la GH en agissant sur 8 acides aminés.
Cette spécificité est donc due à une structure très localisée du récepteur.

Concernant le GHR, ses formes activables et activées sont des homodimères de récepteurs
simples, il est alors nécessaire de faire un regroupement des récepteurs. Les homodimères sont inertes
à la surface de la cellule. Ils comportent deux sites de liaisons à leur partie extracellulaire:
un site 1 de haute affinité ;
un site 2 de basse affinité ;
on appelle le site d’interaction entre les récepteurs se nomme le site 3.
La GH induit un changement de conformation. On observe une rotation relative d’une des
deux chaînes par rapport à l’autre, ce qui va alors entraîner la voie de signalisation.
Le GHR est constitué d’un seul type de chaînes, alors que d’autres sont plus complexes. Il est
possible de trouver des hétérodimères, des hétérotrimères et d’autres choses encore. Il existe au sein de
la superfamille des RCHCI des sous-groupes dans lesquels on trouve une chaîne commune
représentative du sous-groupe, et d’autres chaînes spécifiques pour chaque membre du sous-groupe.
On retrouve ce type d’homodimérisation du récepteur pour les hormones suivantes : la GH,
la PRL, l’EPO, et le G-CSF (un GF spécifique des polynucléaires neutrophiles).
C- Transmission du signal intracellulaire
Le domaine cytoplasmique des récepteurs cytokines est particulier. Il ne comporte pas
d’activité tyrosine-kinase. C’est pourquoi il dispose de kinases associées qui vont assurer la
continuité des voies de signalisation.
Les kinases phosphorylent les acides aminés Tyr, Ser, Thr. La phosphorylation est une
action qui permet d’activer certaines enzymes et de créer des interactions protéine-protéine. Il est
important de rappeler que les tyrosines phosphorylées sont reconnues spécifiquement par des
domaines consensus tels que SH2 et PTB.
Les cascades de signalisations des RCHCI rend compte finalement de 4 fonctions majeures, à
l’échelle cellulaire : la migration, la prolifération, la survie, la différenciation cellulaire.
La continuité du signal se poursuit avec une voie de signalisation spécifique, qui est la voie
JAK/STAT.
Les JAK (Janus kinases) sont des tyrosines-kinases. Il en existe 4 types. Celle qui nous
intéresse principalement est la JAK-2. Les JAK-2 sont liés avec le récepteur via la boîte 1. Le ligand
se fixe au récepteur homodimérisé. La rotation relative des chaînes du récepteur entraîne une mise en
contact des JAK-2. À leurs contact Les JAK se transphosphorylent. Une fois les JAK phosphorylées,
elles phosphorylent le récepteur à sa partie C-terminale, en intracellulaire.
Or, il se trouve que, dans le cytoplasme, il existe des facteurs de transcription nommés STAT
(Signal Transducers and Activators of Transcription). Ces STAT possèdent un domaine SH2 qui
reconnaît les tyrosines phosphorylées.
Les STAT et notamment les STAT 5 viennent fixer avec les phosphotyrosines du récepteur
(ol a déjà été remarqué que les STAT pouvaient venir fixer les JAKs, mais c’est beaucoup moins
fréquent et pas vraiment un cas d’école).
Pendant ce temps, les JAK viennent phosphoryler les STAT sur une tyrosine C-terminale.
Cette tyrosine est capitale au bon fonctionnement de la voie de signalisation. Une mutation de cet
acide aminé pourrait venir impacter la suite des évènements.
Les STAT, après avoir été phosphorylées par les JAK, se détachent du récepteur et se
dimérisent. Le SH2 de l’une lie la phosphotyrosine de l’autre, et vice versa.
 6
7
8
9
10
11
12
13
6
7
8
9
10
11
12
13
1
/
13
100%