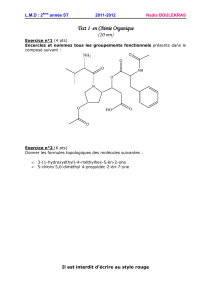
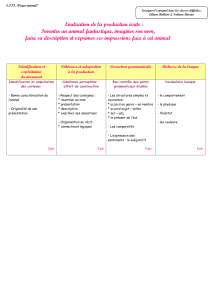
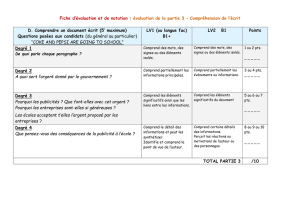
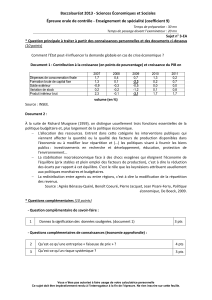
4.3. Les transporteurs secondaires

4.3. Les transporteurs secondaires
4. 3.1. Généralités
- Le transport actif primaire fournit un gradient électrochimique d’ion ou de soluté
utilisé par le transporteur secondaire (primaire = flux d’électrons de la chaîne
respiratoire qui crée le gradient de H+).
- Le transporteur secondaire permet le passage de deux solutés, l’un qui diffuse
spontanément (Na+, H+) et fournit de l’énergie à l’autre qui lui, est transporté
activement.
S/S-
H+/Na+
S/S- H+/Na+
S+ H+/Na+
S+ H+/Na+
Symport Antiport

- Molécules transportées : ions, sucres, aa, peptides, acides carboxyliques,
nucléosides…
- Système de transport très répandu chez les bactéries, 77 familles répertoriées
(selon similarités de séquences)
- La famille la plus importante, MFS (Major Facilitator Superfamilly)
400 à 600 aa, 12, 14, 24 hélices transmembranaires
Un membre bien étudié : La lactose perméase d’
E
.
coli

ext
int
Membrane
Le transport primaire permet de créer et de maintenir le gradient de proton
ext/int = différence de potentiel électrochimique
Le gradient de Na+ est créé par des transporteurs antiport H+/Na+ ou par des
transporteur à ATP
H+ Na+
H+ Na+
Na+ Na+
Na+
Na+
Na+
H+
H+
H+
H+
Membrane
plasmique
(0.5) (150mV)
pH7
pH7.5

4.3.2 Organisation structurale de LacY
- Spécificité pour le lactose et le galactose
- Ratio : 1 proton transporté pour 1 lactose, en symport
- Semble agir à l’état monomérique
- 12 hélices α transmembranaires
- Située dans la membrane cytoplasmique de
E. coli

LacY LacY
H-LacY
H-L-LacY
H+ H+
L L
LacY-H-L
LacY-H
Périplasme Cytoplasme
1
2 34
5
6
4.3.3 Mécanisme proposé du symport
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
1
/
24
100%