Angelman (syndrome d`)

Angelman (syndrome d’)
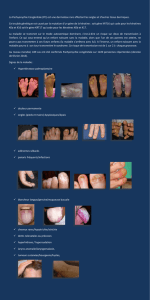
Description clinique de la maladie
Le syndrome d’Angelman (MIM# 105830), également
appelé «syndrome du pantin hilare »(happy puppet
syndrome), aété décrit en 1965 par un pédiatre anglais
et comporte dans sa définition princeps un retard
mental sévère, une ataxie avec démarche raide et mou-
vements saccadés et des traits comportementaux par-
ticuliers de type pantin joyeux.
Le phénotype varie selon l’âge et les patients.
L’histoire naturelle de la maladie débute vers 1an,
période àlaquelle apparaissent le retard des acquisi-
tions motrices, la déficience intellectuelle, l’ataxie asso-
ciée àune hypertonie périphérique donnant àces
enfants lors de l’acquisition de la marche, vers 3–4 ans,
une démarche saccadée. La microcéphalie, constante,
est associée àune dysmorphie faciale.
Le comportement, constant, se caractérise par des rires
fréquents, inadaptés, associés àune hyperexcitabilité,
une hyperactivité avec des troubles du sommeil. Une
épilepsie est présente dans plus de 96 %des cas.
Le diagnostic différentiel est le syndrome de Rett chez
la fille et le retard mental de l’α-thalassémie liée àl’X
chez le garçon.
Le traitement repose sur la kinésithérapie, le traitement
de l’épilepsie et des troubles du sommeil, et la prise en
charge du retard mental.
Épidémiologie et aspect génétique
L’incidence du syndrome d’Angelman est estimée entre
1/12 000 à1/20 000 environ.
Tout comme le syndrome de Prader-Willi, c’est un
modèle de maladie génétique lié au phénomène de
l’empreinte parentale.
Les gènes responsables sont localisés sur le chromo-
some 15, dans la région 15q11-15q12 (gène UBE3A).
Le gène UBE3A code pour une protéine E6-AP, ligase
ubiquitaire, dont le rôle dans les modèles animaux n’est
pas clair. Physiologiquement et àl’état normal, la copie
d’origine maternelle est inactive et seuls les gènes situés
sur le chromosome d’origine paternelle sont actifs.
L’activation des gènes est régulée par un mécanisme de
méthylation différentielle :lesite d’empreinte est situé
2Mbenaval du gène UBE3A.
Cette activation génique est tissu-dépendante pour le
syndrome d’Angelman.
Les mécanismes responsables de ce syndrome sont donc
les mécanismes affectant le site d’empreinte ou le gène
UBE3A lui-même, àsavoir :
•la délétion de la région 15q11-q12 (70 %des cas).
Cette délétion dont l’étendue peut varier contient
dans tous les cas une région commune testée par la
sonde employée pour la technique FISH ;
•une disomie uniparentale paternelle, présence de la
copie d’origine paternelle en double dose (5 %des cas) ;
•la mutation d’empreinte ou la petite délétion du
centre d’empreinte, dont la conséquence est l’absence
de méthylation de la région 15q11/15q12 (5–10 %
des cas) ;
•une mutation du gène UBE3A,dont la fréquence
semble être de l’ordre de 4à5%;
•enfin, pour 5à10 %des patients, le mécanisme molé-
culaire reste inconnu.
Diagnostic moléculaire
L’intérêt d’établir le mécanisme moléculaire réside dans
le conseil génétique et les possibilités de transmission
lors des grossesses suivantes.
Quelle que soit l’anomalie moléculaire en cause, la
résultante est toujours une anomalie de méthylation du
locus. La stratégie du diagnostic biologique est donc
basée sur la vérification de la présence des copies d’ori-
gine maternelle et paternelle par méthyl-PCR.
Cette technique, par sa simplicité, asupplanté les tech-
niques de southern blot avec des sondes sensibles àla
méthylation ou les techniques de PCR-RFLP.
Les ADN extraits sont traités au bisulfite, qui possède
la propriété de transformer les bases cytosines en bases
uracil lorsque celles-ci ne sont pas méthylées (comme
c’est le cas de l’allèle paternel actil lorsqu’il est présent).
L’utilisation d’amorces spécifiques pour chacun des
allèles méthylés ou non permet la reconnaissance de
chacun des brins maternels et paternels.
La présence de l’allèle paternel et de l’allèle maternel
exclut àenviron 85 %lediagnostic de syndrome
d’Angelman, àl’exclusion des réarrangements chromo-
somiques et des mutations du gène UBE3A non détec-
tés par le test. Le nombre de mutations du gène UBE3A
dans le syndrome d’Angelman est élevé. Il n’existe pas
de hot spot.Elles peuvent être de novo ou héritées.
En cas de présence exclusive des copies paternelles (en
simple ou double dose), la confirmation du diagnostic
et la vérification du mécanisme s’effectuent en recher-
chant la délétion par technique FISH en premier lieu
puis par recherche de la disomie uniparentale par étude
des marqueurs microsatellites parentaux comparés au
cas index.

Le syndrome de Prader-Willi est sous-tendu par un
mécanisme moléculaire dans la même région.
☞Disomie uniparentale, Prader-Willi (syndrome de)
(HarveyJ,VoelckelM,Malzac P, Moncla A, Ramsden S, MatthijsG.
Draftbest practiceguidelines formolecularanalysis of Prader Willi and
Angelman syndromes. April2002.
Disponible sur:
http://www.emqn.org /emqn/BestPractice/mainColumnParagraphs/08/
document/PWAS_eu.pdf
JiangYH, Lev-Lehman E, BresslerJ,TsaiTF, Beaudet AL.
GeneticsofAngelman syndrome.
Am JHum Genet 1999 ;65:1-6.
Moncla A.
Le syndrome d’Angelman. Janvier 2000.
Disponible sur:http://www.orpha.net/data/patho/FR/fr-angelman.pdf
1
/
2
100%