Synthèse des acides gras et triglycérides

Synthèse des acides gras et des triglycérides
Introduction :
Cʼest un phénomène très important car les acides gras sont la principale réserve
énergétique. La biosynthèse des ag a lieu dans le foie, le tissu adipeux, et lʼintestin.
Elle est réalisée par des voies métaboliques et dans des compartiments subcellulaires
différents de la dégradation oxydative.
Elle nécessite essentiellement 2 substrats (en plus de lʼATP) : lʼacétyl-CoA et le NADPH.
Dʼou proviennent ces 2 substrats :
- le NADPH est fourni soit par la voie des pentos-phos soit par la réaction catalysée par
lʼenzyme malique
- lʼacétyl-CoA provient essentiellement de la dégradation des glucides. La glucolyse
aboutit au pyruvate qui est tranformé en acCoA dans la mitochondrie. La sortie de
lʼactéyl-CoA se fait de manière indirecte par lʼintermédiaire du citrate qui utilise le
transporteur des tricarboxylates. Dans le cytoplasme, lʼactéyl-CoA est reformé par la
réaction catalysée par la citrate lyase :
1
SYNTHESE
SYNTHESE DES ACIDES GRAS ET TRIGLYCERIDES
DES ACIDES GRAS ET TRIGLYCERIDES
INTRODUCTION
INTRODUCTION
I
I –
–BIOSYNTHESE DES ACIDES GRAS
BIOSYNTHESE DES ACIDES GRAS
II
II –
–BIOSYNTHESE DES TRIGYCERIDES
BIOSYNTHESE DES TRIGYCERIDES
III
III -
-REGULATION DE LA BIOSYNTHESE
REGULATION DE LA BIOSYNTHESE
SYNTHESE
SYNTHESE DES ACIDES GRAS ET TRIGLYCERIDES
DES ACIDES GRAS ET TRIGLYCERIDES
INTRODUCTION
INTRODUCTION
La biosynth
La biosynthè
èse des acides gras a lieu dans le foie, le tissu
se des acides gras a lieu dans le foie, le tissu
adipeux et l
adipeux et l’
’intestin.
intestin.
Elle est r
Elle est ré
éalis
alisé
ée par des voies m
e par des voies mé
étaboliques et dans des
taboliques et dans des
compartiments subcellulaires diff
compartiments subcellulaires diffé
érents de la d
rents de la dé
égradation
gradation
oxydative.
oxydative.
Elle n
Elle né
écessite essentiellement 2 substrats (en plus de l
cessite essentiellement 2 substrats (en plus de l’
’ATP
ATP):
):
l
l’
’ac
acé
étyl
tyl-
-CoA
CoA et le NADPH
et le NADPH
INTRODUCTION (suite)
INTRODUCTION (suite)
D
D’
’o
où
ùproviennent ces 2 substrats:
proviennent ces 2 substrats:
*
* le NADPH
le NADPH est fourni soit par la voie des
est fourni soit par la voie des pentose
pentose-
-phosphates
phosphates,
,
soit par la r
soit par la ré
éaction catalys
action catalysé
ée par l
e par l’
’enzyme malique.
enzyme malique.
*
* l’acétyl-CoA provient essentiellement de la d
provient essentiellement de la dé
égradation des
gradation des
glucides.
glucides.
La glycolyse aboutit au
La glycolyse aboutit au pyruvate
pyruvate qui est transform
qui est transformé
éen
en ac
acé
étyl
tyl-
-
CoA
CoA dans la mitochondrie. La sortie de l
dans la mitochondrie. La sortie de l’
’ac
acé
étyl
tyl-
-CoA
CoA se fait de
se fait de
mani
maniè
ère indirecte par l
re indirecte par l’
’interm
intermé
édiaire du citrate qui utilise le
diaire du citrate qui utilise le
transporteur des
transporteur des tricarboxylates
tricarboxylates.
.
Dans le cytoplasme, l
Dans le cytoplasme, l’
’ac
acé
étyl
tyl-
-CoA
CoA est reform
est reformé
épar la r
par la ré
éaction
action
catalys
catalysé
ée par la citrate
e par la citrate lyase
lyase:
:
Citrate + ATP + CoA-SH + H
2
OOA + Acétyl-CoA + ADP + Pi
La synthèse des AG requiert la coopération de plusieurs voies
métaboliques situées dans des compartiments cellulaires différents.
NB: l’acétyl-CoA stimule la citrate synthase
La synthèse des AG requiert la coopération de plusieurs voies métaboliques situées dans
des compartiments cellulaires différents. La source des unités dicarbonée est le glucose
pour synthétiser les ag, il est transformé en pyruvate au niveau de la glycolyse, le pyruvate
traverse la m mitochondriale, il peut donner de lʼaccoa de lʼoxaloatétate, qui peuvent se
combiner au niveau de la première étape du cycle de Krebs pour donner du citrate. Si on
a un potentiel élevé, on a de lʼATP en grande quté et les enzymes sont inhibés, le cycle de
Krebs est ralenti, le citrate sera donc utilisé pour la synthèse des ag, il va sortir de la
mitonchondrie par son transporteur : les tricarboxylates. Lʼactéyl coa est un stimulateur de
la citrate synthase, la concentration élevée en actéyl co permet la synthèse du citrate mais
le cycle ne fonctionne pas. On a un passage du citrate au nivau du cytosome grace a la
citrate lyase. Lʼoxaloacétate sera transformé en malate ou citrate pour le NADPH.
1
SYNTHESE
SYNTHESE DES ACIDES GRAS ET TRIGLYCERIDES
DES ACIDES GRAS ET TRIGLYCERIDES
INTRODUCTION
INTRODUCTION
I
I –
–BIOSYNTHESE DES ACIDES GRAS
BIOSYNTHESE DES ACIDES GRAS
II
II –
–BIOSYNTHESE DES TRIGYCERIDES
BIOSYNTHESE DES TRIGYCERIDES
III
III -
-REGULATION DE LA BIOSYNTHESE
REGULATION DE LA BIOSYNTHESE
SYNTHESE
SYNTHESE DES ACIDES GRAS ET TRIGLYCERIDES
DES ACIDES GRAS ET TRIGLYCERIDES
INTRODUCTION
INTRODUCTION
La biosynth
La biosynthè
èse des acides gras a lieu dans le foie, le tissu
se des acides gras a lieu dans le foie, le tissu
adipeux et l
adipeux et l’
’intestin.
intestin.
Elle est r
Elle est ré
éalis
alisé
ée par des voies m
e par des voies mé
étaboliques et dans des
taboliques et dans des
compartiments subcellulaires diff
compartiments subcellulaires diffé
érents de la d
rents de la dé
égradation
gradation
oxydative.
oxydative.
Elle n
Elle né
écessite essentiellement 2 substrats (en plus de l
cessite essentiellement 2 substrats (en plus de l’
’ATP
ATP):
):
l
l’
’ac
acé
étyl
tyl-
-CoA
CoA et le NADPH
et le NADPH
INTRODUCTION (suite)
INTRODUCTION (suite)
D
D’
’o
où
ùproviennent ces 2 substrats:
proviennent ces 2 substrats:
*
* le NADPH
le NADPH est fourni soit par la voie des
est fourni soit par la voie des pentose
pentose-
-phosphates
phosphates,
,
soit par la r
soit par la ré
éaction catalys
action catalysé
ée par l
e par l’
’enzyme malique.
enzyme malique.
*
* l’acétyl-CoA provient essentiellement de la d
provient essentiellement de la dé
égradation des
gradation des
glucides.
glucides.
La glycolyse aboutit au
La glycolyse aboutit au pyruvate
pyruvate qui est transform
qui est transformé
éen
en ac
acé
étyl
tyl-
-
CoA
CoA dans la mitochondrie. La sortie de l
dans la mitochondrie. La sortie de l’
’ac
acé
étyl
tyl-
-CoA
CoA se fait de
se fait de
mani
maniè
ère indirecte par l
re indirecte par l’
’interm
intermé
édiaire du citrate qui utilise le
diaire du citrate qui utilise le
transporteur des
transporteur des tricarboxylates
tricarboxylates.
.
Dans le cytoplasme, l
Dans le cytoplasme, l’
’ac
acé
étyl
tyl-
-CoA
CoA est reform
est reformé
épar la r
par la ré
éaction
action
catalys
catalysé
ée par la citrate
e par la citrate lyase
lyase:
:
Citrate + ATP + CoA-SH + H
2
OOA + Acétyl-CoA + ADP + Pi
La synthèse des AG requiert la coopération de plusieurs voies
métaboliques situées dans des compartiments cellulaires différents.
NB: l’acétyl-CoA stimule la citrate synthase
NB : lʼacétyl-CoA stimule la citrate synthase

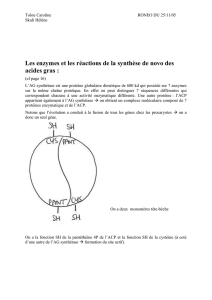
I- Biosynthèse des acides gras
1- Formation du malonyl-CoA
Le donnneur activé dʼunités dicarbonées est le malonyl-CoA. Réaction des 2 acides gras
avec les 2 coA défavorisée donc : Réaction de Décarboxylation . Enzyme : acétyl-CoA
carboxylase.
2
SYNTHESE
SYNTHESE DES ACIDES GRAS ET TRIGLYCERIDES
DES ACIDES GRAS ET TRIGLYCERIDES
INTRODUCTION
INTRODUCTION
I
I –
–BIOSYNTHESE DES ACIDES GRAS
BIOSYNTHESE DES ACIDES GRAS
1
1°
°Formation du
Formation du Malonyl
Malonyl-
-CoA
CoA
2
2°
°Synth
Synthè
èse de l
se de l’
’Ac
Ac Gras par le complexe AGS
Gras par le complexe AGS
II
II –
–BIOSYNTHESE DES TRIGYCERIDES
BIOSYNTHESE DES TRIGYCERIDES
III
III -
-REGULATION DE LA BIOSYNTHESE
REGULATION DE LA BIOSYNTHESE
-
OOC CH2CS-CoA
O
Malonyl
Malonyl-
-CoA
CoA
+ ADP + Pi + H+
CH3CS-CoA
O
Ac
Acé
étyl
tyl-
-CoA
CoA
+ HCO3
-
+ ATP
Ac
Acé
étyl
tyl-
-CoA
CoA Carboxylase
Carboxylase
Etape d’engagement irréversible:
rôle-clé dans la régulation de la biosynthèse
Le donneur activé d’unités dicarbonées est le malonyl-CoA
Ac
Acé
étyl
tyl-
-CoA
CoA Carboxylase
Carboxylase
ATP ADP + Pi
CO
2
E - Biotine E - Biotine
~
COO
-
Malonyl
Malonyl-
-CoA
CoA Ac
Acé
étyl
tyl-
-CoA
CoA
Liaison
Liaison à
àun
un
r
ré
ésidu Lys
sidu Lys
Carboxybiotinyl
Carboxybiotinyl
enzyme
enzyme
SYNTHESE
SYNTHESE DES ACIDES GRAS ET TRIGLYCERIDES
DES ACIDES GRAS ET TRIGLYCERIDES
INTRODUCTION
INTRODUCTION
I
I –
–BIOSYNTHESE DES ACIDES GRAS
BIOSYNTHESE DES ACIDES GRAS
1
1°
°Formation du
Formation du Malonyl
Malonyl-
-CoA
CoA
2
2°
°Synth
Synthè
èse de l
se de l’
’Ac
Ac Gras par le complexe AGS
Gras par le complexe AGS
II
II –
–BIOSYNTHESE DES TRIGYCERIDES
BIOSYNTHESE DES TRIGYCERIDES
III
III -
-REGULATION DE LA BIOSYNTHESE
REGULATION DE LA BIOSYNTHESE
Etape dʼengagement irréversible : role-clé dans la régulation de la biosynthèse.
Acétyl-CoA carboxylase
2
SYNTHESE
SYNTHESE DES ACIDES GRAS ET TRIGLYCERIDES
DES ACIDES GRAS ET TRIGLYCERIDES
INTRODUCTION
INTRODUCTION
I
I –
–BIOSYNTHESE DES ACIDES GRAS
BIOSYNTHESE DES ACIDES GRAS
1
1°
°Formation du
Formation du Malonyl
Malonyl-
-CoA
CoA
2
2°
°Synth
Synthè
èse de l
se de l’
’Ac
Ac Gras par le complexe AGS
Gras par le complexe AGS
II
II –
–BIOSYNTHESE DES TRIGYCERIDES
BIOSYNTHESE DES TRIGYCERIDES
III
III -
-REGULATION DE LA BIOSYNTHESE
REGULATION DE LA BIOSYNTHESE
-
OOC CH2CS-CoA
O
Malonyl
Malonyl-
-CoA
CoA
+ ADP + Pi + H+
CH3CS-CoA
O
Ac
Acé
étyl
tyl-
-CoA
CoA
+ HCO3
-
+ ATP
Ac
Acé
étyl
tyl-
-CoA
CoA Carboxylase
Carboxylase
Etape d’engagement irréversible:
rôle-clé dans la régulation de la biosynthèse
Le donneur activé d’unités dicarbonées est le malonyl-CoA
Ac
Acé
étyl
tyl-
-CoA
CoA Carboxylase
Carboxylase
ATP ADP + Pi
CO
2
E - Biotine E - Biotine
~
COO
-
Malonyl
Malonyl-
-CoA
CoA Ac
Acé
étyl
tyl-
-CoA
CoA
Liaison
Liaison à
àun
un
r
ré
ésidu Lys
sidu Lys
Carboxybiotinyl
Carboxybiotinyl
enzyme
enzyme
SYNTHESE
SYNTHESE DES ACIDES GRAS ET TRIGLYCERIDES
DES ACIDES GRAS ET TRIGLYCERIDES
INTRODUCTION
INTRODUCTION
I
I –
–BIOSYNTHESE DES ACIDES GRAS
BIOSYNTHESE DES ACIDES GRAS
1
1°
°Formation du
Formation du Malonyl
Malonyl-
-CoA
CoA
2
2°
°Synth
Synthè
èse de l
se de l’
’Ac
Ac Gras par le complexe AGS
Gras par le complexe AGS
II
II –
–BIOSYNTHESE DES TRIGYCERIDES
BIOSYNTHESE DES TRIGYCERIDES
III
III -
-REGULATION DE LA BIOSYNTHESE
REGULATION DE LA BIOSYNTHESE
La biotine est liée a lʼenzyme, grace a lʼATP : carbxylation de la biotine et ensuite transfert
sur lʼactéyl CoA pour donner le malonyl CoA. Cʼest lʼétape dʼengagement et de régulation
de la synthèse des ag.
2- Synthèse de lʼag par le complexe AGsynthase
Les intermédiaires de la synthèse des ag sont attachés a une protéine de trasnprot dʼacyle
ou ACP.
La synthèse des ag consiste en plusieurs séries de réaction :
- condensation
- réduction
- déshydratation
- réduction
Les différentes activités enzymatiques sont réunies en une seule chaine polypeptidique
appelée A G synthase ou AGS :
- domaine 1 : unité dʼentrée et de condensation des substrats. Un tour de cyle on a un ag
a 4 carbone : le butiryl coA, puis nouveau cycle grace a un nouveau malonyl CoA (6C) ....
- domaine 2 : unité de réduction
- domaine 3 : unité de libération de lʼag : lorsque lʼon a complètement finie la synthèse
(C16) en général on a une enzyme qui permet de libérer lʼag de lʼACP (fait partie de
lʼAGS)

Structure de lʼAGS de mammifères : la forme active est un dimère
3
Synthèse des Acides Gras
•Les intermédiaires de la synthèse des AG sont attachés à une protéine
de transport d’acyles ou ACP
•La synthèse des acides gras consiste en plusieurs séries de réaction:
-condensation
-réduction
-deshydratation
-réduction
•Les différentes activités enzymatiques sont réunies en une seule
chaîne polypeptidique appelée Acide Gras Synthase ou AGS:
-domaine 1: unitéd’entrée et de condensation des substrats
-domaine 2: unitéde réduction
-domaine 3: unitéde libération de l’acide gras
Structure de l’AGS de mammifères: la forme active est un dimère
CSCoA
O
CH
3
C
O
SCoA
Malonyl
Malonyl-
-CoA
CoA Ac
Acé
étyl
tyl-
-CoA
CoA
-
OOC-CH
2
AT
AT
CSACP
O
CH
3
C
O
SCys-
Malonyl
Malonyl-
-ACP
ACP Ac
Acé
étyl
tyl-
-Enz
Enz (CAS)
(CAS)
-
OOC-CH
2
MT
MT
Transfert de l’acétyle et du malonyle sur l’AGS
HS-CoA
CH
3
CSACP
O
CH
2
C
O
Ac
Acé
éto
to-
-ac
acé
étyl
tyl-
-ACP
ACP
(
(Ac
Acé
éto
to-
-acyl
acyl-
-ACP
ACP)
)
CSACP
O
+ CH
3
C
O
SACP
R
Ré
éduction (CAR)
duction (CAR)
Malonyl
Malonyl-
-ACP
ACP Ac
Acé
étyl
tyl-
-S
S-
-Enzyme
Enzyme
-
OOC-CH
2
CH
3
C C C S ACP
O
H
HH
OH
3
32
21
1
NADPH
D-3-OH-butyryl-ACP
(D-3-OH-acyl-ACP)
NADP+
Condensation (CAS)
Condensation (CAS)
CO2
3 acitivités enzymatiques :
- domaine 1 : AT, MT et CAS : on a le malonyl coa qui va se positionner sur lʼACP grace
au résidu SH (la malonyl transférase), on a lʼacétyl co grace a lʼactéyl transférase au
niveau de la CAS par un groupement SH, cette protéine va transférer lʼacétyl sur le
malonyl.
Transfert de lʼacétyle et du malonyle sur lʼAGS
3
Synthèse des Acides Gras
•Les intermédiaires de la synthèse des AG sont attachés à une protéine
de transport d’acyles ou ACP
•La synthèse des acides gras consiste en plusieurs séries de réaction:
-condensation
-réduction
-deshydratation
-réduction
•Les différentes activités enzymatiques sont réunies en une seule
chaîne polypeptidique appelée Acide Gras Synthase ou AGS:
-domaine 1: unitéd’entrée et de condensation des substrats
-domaine 2: unitéde réduction
-domaine 3: unitéde libération de l’acide gras
Structure de l’AGS de mammifères: la forme active est un dimère
CSCoA
O
CH
3
C
O
SCoA
Malonyl
Malonyl-
-CoA
CoA Ac
Acé
étyl
tyl-
-CoA
CoA
-
OOC-CH
2
AT
AT
CSACP
O
CH
3
C
O
SCys-
Malonyl
Malonyl-
-ACP
ACP Ac
Acé
étyl
tyl-
-Enz
Enz (CAS)
(CAS)
-
OOC-CH
2
MT
MT
Transfert de l’acétyle et du malonyle sur l’AGS
HS-CoA
CH
3
CSACP
O
CH
2
C
O
Ac
Acé
éto
to-
-ac
acé
étyl
tyl-
-ACP
ACP
(
(Ac
Acé
éto
to-
-acyl
acyl-
-ACP
ACP)
)
CSACP
O
+ CH
3
C
O
SACP
R
Ré
éduction (CAR)
duction (CAR)
Malonyl
Malonyl-
-ACP
ACP Ac
Acé
étyl
tyl-
-S
S-
-Enzyme
Enzyme
-
OOC-CH
2
CH
3
C C C S ACP
O
H
HH
OH
3
32
21
1
NADPH
D-3-OH-butyryl-ACP
(D-3-OH-acyl-ACP)
NADP+
Condensation (CAS)
Condensation (CAS)
CO2
Si on fait plusieurs tours on a lʼacéto acyl ACP
- domaine 2 : CAR, HAD et ER : réduction déshydratation et réduction

3
Synthèse des Acides Gras
•Les intermédiaires de la synthèse des AG sont attachés à une protéine
de transport d’acyles ou ACP
•La synthèse des acides gras consiste en plusieurs séries de réaction:
-condensation
-réduction
-deshydratation
-réduction
•Les différentes activités enzymatiques sont réunies en une seule
chaîne polypeptidique appelée Acide Gras Synthase ou AGS:
-domaine 1: unitéd’entrée et de condensation des substrats
-domaine 2: unitéde réduction
-domaine 3: unitéde libération de l’acide gras
Structure de l’AGS de mammifères: la forme active est un dimère
CSCoA
O
CH
3
C
O
SCoA
Malonyl
Malonyl-
-CoA
CoA Ac
Acé
étyl
tyl-
-CoA
CoA
-
OOC-CH
2
AT
AT
CSACP
O
CH
3
C
O
SCys-
Malonyl
Malonyl-
-ACP
ACP Ac
Acé
étyl
tyl-
-Enz
Enz (CAS)
(CAS)
-
OOC-CH
2
MT
MT
Transfert de l’acétyle et du malonyle sur l’AGS
HS-CoA
CH
3
CSACP
O
CH
2
C
O
Ac
Acé
éto
to-
-ac
acé
étyl
tyl-
-ACP
ACP
(
(Ac
Acé
éto
to-
-acyl
acyl-
-ACP
ACP)
)
CSACP
O
+ CH
3
C
O
SACP
R
Ré
éduction (CAR)
duction (CAR)
Malonyl
Malonyl-
-ACP
ACP Ac
Acé
étyl
tyl-
-S
S-
-Enzyme
Enzyme
-
OOC-CH
2
CH
3
C C C S ACP
O
H
HH
OH
3
32
21
1
NADPH
D-3-OH-butyryl-ACP
(D-3-OH-acyl-ACP)
NADP+
Condensation (CAS)
Condensation (CAS)
CO2
4
CH
3
C C C S ACP
O
H
H
Crotonyl
Crotonyl-
-ACP
ACP
(!
(!(!
(!
(!
(!(!
(!2
2-
-Enoyl
Enoyl-
-ACP)
ACP)
2
21
1
R
Ré
éduction (ER)
duction (ER)
NADPH
NADP+
CH
3
CH
2
CH
2
C S ACP
O
Butyryl-ACP
(Acyl-ACP)
CH
3
C C C S ACP
O
H
HH
OH
3
32
21
1
D-3-OH-butyryl-ACP
(D-3-OH-acyl-ACP)
Deshydratation
Deshydratation (HAD)
(HAD)
Addition de 2 carbones
àchaque cycle
Malonyl-CoA
H
2
O
Equation globale de la synth
Equation globale de la synthè
èse du palmitate
se du palmitate
8 Acétyl-CoA + 7 ATP + 14 NADPH + 6 H
+
Palmitate + 8 CoASH + 14 NADP
+
+6 H
2
O + 7 ADP + 7 Pi
Acétyl-CoA + 7 Malonyl-CoA + 14 NADPH + 20 H
+
Palmitate + 8 CoASH + 14 NADP
+
+6 H
2
O + 7 CO
2
Synthèse du malonyl-CoA:
7Acétyl-CoA + 7 CO
2
+ 7 ATP 7 Malonyl-CoA + 7 ADP + 7 Pi + 14H
+
Synthèse du palmitate à partir des acétyl-CoA:
Synthèse du palmitate à partir de l’acétyl-CoA et du malonyl-CoA:
SYNTHESE
SYNTHESE DES ACIDES GRAS ET TRIGLYCERIDES
DES ACIDES GRAS ET TRIGLYCERIDES
INTRODUCTION
INTRODUCTION
I
I –
–BIOSYNTHESE DES ACIDES GRAS
BIOSYNTHESE DES ACIDES GRAS
II
II –
–BIOSYNTHESE DES TRIGYCERIDES
BIOSYNTHESE DES TRIGYCERIDES
III
III -
-REGULATION DE LA BIOSYNTHESE: Rôle
REGULATION DE LA BIOSYNTHESE: Rôle-
-cl
clé
éde
de
Les Acides gras sont mis en réserve dans le tissu adipeux
sous forme de triglycérides
R
2
C
O
O
CH
2
CH
CH
2
C
O
R
1
O
C
O
R
3
O
Les acides gras proviennent :
-soit d’une synthèse endogène (ce que l’on vient de voir)
-soit de l’alimentation : AG exogènes, majoritaires (alimentation équilibrée)
Le glycérol, sous forme de glycérol phosphate vient de la glycolyse
(par réduction de la dihydroxyacétone phosphate)
L’estérification des 3 fonctions alcool du glycérol par 3 acides gras
aboutit à la formation d’un triglycéride :
Principal site de synthèse et de stockage des TG: cytoplasme des adipocytes
: Déshydratation : départ dʼune
molécule dʼeau.
Cycle jusquʼau C16
- domaine 3 : PTE : elle va repérer la longueur de la chaine de lʼag, et va agir de manière
a libérer lʼag sous forme libre de palmitate, lʼenzyme est une thiolase ou thioestérase
Equation globale de la synthèse du palmitate
4
CH
3
C C C S ACP
O
H
H
Crotonyl
Crotonyl-
-ACP
ACP
(!
(!(!
(!
(!
(!(!
(!2
2-
-Enoyl
Enoyl-
-ACP)
ACP)
2
21
1
R
Ré
éduction (ER)
duction (ER)
NADPH
NADP+
CH
3
CH
2
CH
2
C S ACP
O
Butyryl-ACP
(Acyl-ACP)
CH
3
C C C S ACP
O
H
HH
OH
3
32
21
1
D-3-OH-butyryl-ACP
(D-3-OH-acyl-ACP)
Deshydratation
Deshydratation (HAD)
(HAD)
Addition de 2 carbones
àchaque cycle
Malonyl-CoA
H
2
O
Equation globale de la synth
Equation globale de la synthè
èse du palmitate
se du palmitate
8 Acétyl-CoA + 7 ATP + 14 NADPH + 6 H
+
Palmitate + 8 CoASH + 14 NADP
+
+6 H
2
O + 7 ADP + 7 Pi
Acétyl-CoA + 7 Malonyl-CoA + 14 NADPH + 20 H
+
Palmitate + 8 CoASH + 14 NADP
+
+6 H
2
O + 7 CO
2
Synthèse du malonyl-CoA:
7Acétyl-CoA + 7 CO
2
+ 7 ATP 7 Malonyl-CoA + 7 ADP + 7 Pi + 14H
+
Synthèse du palmitate à partir des acétyl-CoA:
Synthèse du palmitate à partir de l’acétyl-CoA et du malonyl-CoA:
SYNTHESE
SYNTHESE DES ACIDES GRAS ET TRIGLYCERIDES
DES ACIDES GRAS ET TRIGLYCERIDES
INTRODUCTION
INTRODUCTION
I
I –
–BIOSYNTHESE DES ACIDES GRAS
BIOSYNTHESE DES ACIDES GRAS
II
II –
–BIOSYNTHESE DES TRIGYCERIDES
BIOSYNTHESE DES TRIGYCERIDES
III
III -
-REGULATION DE LA BIOSYNTHESE: Rôle
REGULATION DE LA BIOSYNTHESE: Rôle-
-cl
clé
éde
de
Les Acides gras sont mis en réserve dans le tissu adipeux
sous forme de triglycérides
R
2
C
O
O
CH
2
CH
CH
2
C
O
R
1
O
C
O
R
3
O
Les acides gras proviennent :
-soit d’une synthèse endogène (ce que l’on vient de voir)
-soit de l’alimentation : AG exogènes, majoritaires (alimentation équilibrée)
Le glycérol, sous forme de glycérol phosphate vient de la glycolyse
(par réduction de la dihydroxyacétone phosphate)
L’estérification des 3 fonctions alcool du glycérol par 3 acides gras
aboutit à la formation d’un triglycéride :
Principal site de synthèse et de stockage des TG: cytoplasme des adipocytes

II- Biosynthèse des triglycérides
Les ag sont mis en réserve dans le tissu adipeux sous forme de triglycérides.
Les ag proviennent :
- soit dʼune synthèse endogène (ce que lʼon vient de voir)
- soit de lʼalimentation : ag exogènes, majoritaire (alimentation équilibrée)
Le glycérol sous forme de glycérilphosphate vient de la glycoluse (par réduction de la
dihydroxyacétone phosphate)
Lʼestérification des 3 fonctions alcool du glycérol par 3 ag aboutit a la formation dʼun
triglycéride :
4
CH
3
C C C S ACP
O
H
H
Crotonyl
Crotonyl-
-ACP
ACP
(!
(!(!
(!
(!
(!(!
(!2
2-
-Enoyl
Enoyl-
-ACP)
ACP)
2
21
1
R
Ré
éduction (ER)
duction (ER)
NADPH
NADP+
CH
3
CH
2
CH
2
C S ACP
O
Butyryl-ACP
(Acyl-ACP)
CH
3
C C C S ACP
O
H
HH
OH
3
32
21
1
D-3-OH-butyryl-ACP
(D-3-OH-acyl-ACP)
Deshydratation
Deshydratation (HAD)
(HAD)
Addition de 2 carbones
àchaque cycle
Malonyl-CoA
H
2
O
Equation globale de la synth
Equation globale de la synthè
èse du palmitate
se du palmitate
8 Acétyl-CoA + 7 ATP + 14 NADPH + 6 H
+
Palmitate + 8 CoASH + 14 NADP
+
+6 H
2
O + 7 ADP + 7 Pi
Acétyl-CoA + 7 Malonyl-CoA + 14 NADPH + 20 H
+
Palmitate + 8 CoASH + 14 NADP
+
+6 H
2
O + 7 CO
2
Synthèse du malonyl-CoA:
7Acétyl-CoA + 7 CO
2
+ 7 ATP 7 Malonyl-CoA + 7 ADP + 7 Pi + 14H
+
Synthèse du palmitate à partir des acétyl-CoA:
Synthèse du palmitate à partir de l’acétyl-CoA et du malonyl-CoA:
SYNTHESE
SYNTHESE DES ACIDES GRAS ET TRIGLYCERIDES
DES ACIDES GRAS ET TRIGLYCERIDES
INTRODUCTION
INTRODUCTION
I
I –
–BIOSYNTHESE DES ACIDES GRAS
BIOSYNTHESE DES ACIDES GRAS
II
II –
–BIOSYNTHESE DES TRIGYCERIDES
BIOSYNTHESE DES TRIGYCERIDES
III
III -
-REGULATION DE LA BIOSYNTHESE: Rôle
REGULATION DE LA BIOSYNTHESE: Rôle-
-cl
clé
éde
de
Les Acides gras sont mis en réserve dans le tissu adipeux
sous forme de triglycérides
R
2
C
O
O
CH
2
CH
CH
2
C
O
R
1
O
C
O
R
3
O
Les acides gras proviennent :
-soit d’une synthèse endogène (ce que l’on vient de voir)
-soit de l’alimentation : AG exogènes, majoritaires (alimentation équilibrée)
Le glycérol, sous forme de glycérol phosphate vient de la glycolyse
(par réduction de la dihydroxyacétone phosphate)
L’estérification des 3 fonctions alcool du glycérol par 3 acides gras
aboutit à la formation d’un triglycéride :
Principal site de synthèse et de stockage des TG: cytoplasme des adipocytes
Principal site de synthèse et de stockage sont les adipocytes
III- Régulation de la biosynthèse
En relation avec lʼacétyl CoA carboxylase qui a un role clé
La biosynthèse des ag est régulée a court terme par la disponibilité des substrats et par
des régulations enzymatiques (phosphorylation/dé) (si on a un excès de glucide, on a un
excès dʼacétyl coa => donc synthèse dʼAG)
1- Régulation allostéroique
Lʼacétyl CoA carboxylase est :
- activée par le citrate (polymérisation de lʼenzyme) : potentiel énergétique très élevé de la
cellule : polymérisation de lʼenzyme qui devient beaucoup plus active
- inhibiée par le produit final acyl-CoA (palmityl CoA)
2- Régulation par phosphorylation/dé
- Sous dépendance hormonale : insuline, adrénaline, glucagon
- Dans les situations de carence énergétiques : role de la protéine kinase AMP-
dépendante ou AMPL
Inactive Ph, et Active déph. Lʼadrénaline et le glucagon active la protéine kinase a qui va
phos lʼacétyl co a carboxylase et qui transforme lʼenzyme sous une forme inactive on a
une inhibition de la synthèse des ag.
Autre régulation de type métabolique par lʼAMP, grace a lʼAMPK (kinase). Si on a une forte
concentration en AMP (peu dʼATP) on a une régulation allostérique qui aboutit a
lʼinactivation de lʼenzyme et si on a besoin dʼénergie on fait la dégradation de lʼacétyl coA.
5
SYNTHESE
SYNTHESE DES ACIDES GRAS ET TRIGLYCERIDES
DES ACIDES GRAS ET TRIGLYCERIDES
INTRODUCTION
INTRODUCTION
I
I –
–BIOSYNTHESE DES ACIDES GRAS
BIOSYNTHESE DES ACIDES GRAS
II
II –
–BIOSYNTHESE DES TRIGYCERIDES
BIOSYNTHESE DES TRIGYCERIDES
III
III -
-REGULATION DE LA BIOSYNTHESE
REGULATION DE LA BIOSYNTHESE:
:
Rôle
Rôle-
-cl
clé
éde l
de l’
’ac
acé
étyl
tyl-
-CoA
CoA carboxylase
carboxylase
1
1°
°R
Ré
égulation allost
gulation allosté
érique
rique
2
2°
°R
Ré
égulation hormonale
gulation hormonale
III
III -
-REGULATION DE LA BIOSYNTHESE:
REGULATION DE LA BIOSYNTHESE:
La biosynth
La biosynthè
èse des acides gras est r
se des acides gras est ré
égul
gulé
ée
e à
àcourt terme par la
court terme par la
disponibilit
disponibilité
édes substrats et par des r
des substrats et par des ré
égulations enzymatiques
gulations enzymatiques
(phosphorylation/d
(phosphorylation/dé
éphosphorylation)
phosphorylation)
Rôle
Rôle-
-cl
clé
éde
de l’acétyl-CoA carboxylase (ACC):
1
1°
°R
Ré
égulation allost
gulation allosté
érique:
rique:
l
l’
’ac
acé
étyl
tyl-
-CoA
CoA carboxylase est:
carboxylase est:
-activée par le citrate (polymérisation de l’enzyme)
-inhibée par le produit final acyl-CoA(palmityl-CoA)
2° Régulation par phosphorylation/déphosphorylation
-sous dépendance hormonale: insuline, adrénaline, glucagon
-dans les situations de carence énergétique: rôle de la protéine
kinase AMP-dépendante ou AMPK
ADP
ADP
ATP
ATP
inactive
inactive
ACC
ACC
P
P
active
active
PP2A
PP2A
PKA
PKA
H
H
2
2
O
O
Pi
Pi
ACC
ACC
AMPK
AMPK
Adrénaline
Glucagon
Insuline
+
+
Conclusion: l’insuline stimule la synthèse des acides gras et des triglycérides,
tandis que le glucagon et l’adrénaline ont l’effet opposé
AMP
AMP
+
R
Ré
éserves Energ
serves Energé
étiques
tiques
Homme de 70 kg
kg
kg kJ
kJ
Métabolisme basal = 8 000 kJ/jour (2000 kcal/jour)
Consommation chez un sportif: jusqu’à 12 000 kJ/jour
Survie théorique en cas de grève de la faim !80 jours
Triglycérides (tissu adipeux) 15 560 000
Protéines musculaires 6 100 000
Glycogène musculaire 0,300 5 000
Glycogène hépatique 0,100 1 700
 6
6
1
/
6
100%