Enzymologie
Cinétique enzymatique
E + S ES EP E + P
Sa cinétique est influencée par l’enzyme, le substrat et l’environnement (pH, température,
ions, coenzymes et autres composés). Pour mesurer la vitesse de réaction transformant S en
P catalysée (= diminue l’énergie d’activation donc augmente la vitesse) par l’enzyme E, on a
P = f(t), donc la vitesse de la réaction est : activité enzymatique (AE) v = -
=
kcat [ES] (mol. ), v est tangente à la courbe et varie au cours du temps
1 : phase très brève (invisible),
liaison rapide et réversible E+S,
ES augmente et v est
croissante
2 : toutes les enzymes sont sous
forme ES, ES constante, v
maximale et constante tant que
S >>> P
3 : EP et P augmentent, la
réaction inverse PS
commence à se produire, v
diminue
4 : vitesse réaction directe =
vitesse réaction indirecte, on a
donc l’équilibre, S et P constantes, v nulle et constante
En enzymologie on étudie toujours la phase stationnaire = « conditions de vitesse initiale »,
la vitesse initiale () est :
- La vitesse la plus grande de toutes les vitesses mesurables
- a une variation linéaire
- c’est l’efficacité catalytique maximale
1- Influence de la concentration en enzyme
Conditions pour l’étude :

- E variable, tous les autres paramètres sont constants
- S >>> E
- Mesure pour différentes E
On observe : quand E augmente, la vitesse augmente (tangente plus pentue), on a qui
varie linéairement avec E
2 - Influence du pH, température
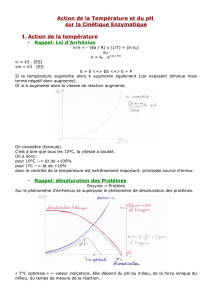
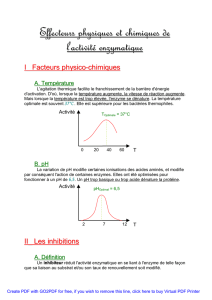
Le pH influe sur l’état d’ionisation des acides aminés, un certain pH peut dénaturer l’enzyme
ou affecter la vitesse de formation ou de dissociation de ES, c’est pourquoi il y a un pH
optimum caractéristique de chaque enzyme.
Selon la loi d’Arrhénius, lorsque l’on augmente la température on accélère la vitesse de la
réaction puis au-delà d’un seuil la température dénature progressivement l’enzyme, c’est
pourquoi il y a une température d’inactivation spécifique de chaque enzyme ; enzymes
biologiques : température optimale de 37C.
3 - Influence de la concentration en substrat : équation de Michaelis-
Menten
Conditions :
- S variable, tous les autres paramètres constants
- S >>> E
- Mesure de pour chaque S
On observe : quand S augmente, augmente ; on a qui donne une hyperbole
équilatère, si S + alors , si S alors 0, si =
alors on a
P = f(t)
Détermination des 2 paramètres cinétiques de l’enzyme :
- Vitesse initiale maximale : activité enzymatique maximale lorsque l’enzyme est
saturée en substrat ( )

- Constante de Michaelis-Menten : concentration de S qui permet d’avoir
- Etape 1 : formation réversible du complexe ES, état pré-stationnaire
- Etape 2 : toutes les enzymes sont complexées, c’est l’état stationnaire : ES
constante et S >>> P, réaction inverse P S est négligée, constante
catalytique (= constante de vitesse de la réaction enzymatique)
avec
Avec
=
en mol. :
- Caractéristique fondamentale du couple E/S
- Ne dépend pas de la concentration en enzyme
- Indicateur de l’affinité de E pour S
- Assimilée à du complexe ES
Avec la constante catalytique = nombre de turn-over = nombre de molécules de
S transformées / sec / molécule de E
et sont des constantes de premier ordre (, est une constante du second
ordre (mol/L/min)
Avec (= ) en mol. qui est la maximale, dépend
de E
Problème : il est difficile d’exploiter la représentation en hyperbole de Michaelis-Menten, on
va donc la linéariser.
Représentation de Lineweaver et Burk
Transformation de Lineweaver et Burk (doubles inverses) :
État pré-stationnaire
État stationnaire
est négligé

4 - Mesure de l’activité enzymatique
Unités de mesure
- UI : unité internationale = quantité d’enzyme transformant 1 mole de substrat par
minute (mol/min) dans les conditions standard (température et pH)
1 UI = 16,66 nkatal
1 nkatal = 60. UI
- Katal (kat) : unité SI = quantité d’enzyme transformant 1 mole de substrat par
seconde (mol/sec)
- Quantité catalytique = en UI = quantité d’enzyme dans un milieu réactionnel
capable de réagir avec le substrat présent dans le milieu
- Concentration catalytique =
en UI/L ou UI/g = quantité
analytique ramenée à 1 g ou 1 L de solution d’enzyme
- Concentration catalytique spécifique =
en UI/mg de protéines
- Activité molaire spécifique ou nombre de rotation (turn-over number) = en
= AS max x MM enzyme =
nombre de molécules de substrat
transformées / sec / molécule d’enzyme
- Activité spécifique (AS) =
(utilisation pour le suivi de purification d’une
enzyme) = activité enzymatique par unité de masse de protéines totales (UI/mg) ou
par volume de solution d’enzyme (UI/L), traduit le degré de pureté de l’enzyme
AS = activité enzymatique / masse totale de protéines
- Indice de purification (sans unité) = degré d’enrichissement en enzyme après
purification par rapport à l’extrait de départ
Enrichissement = AS de la fraction / AS de l’extrait de départ
- Rendement = pourcentage d’enzyme récupéré par rapport à la quantité de départ
R = (activité totale extrait final / activité totale extrait initial) x 100
5 - Influence des inhibiteurs
Les effecteurs sont des molécules de natures diverses qui interfèrent avec une étape de la
catalyse :
- Soit en l’accélérant : activateurs
- Soit en la ralentissant : inhibiteurs (réversibles ou irréversibles)
Ils sont très utiles pour la compréhension des mécanismes enzymatiques et en
thérapeutique.
a) Inhibitions « irréversibles »
En cas de liaison covalente de l’inhibiteur à l’enzyme on parle d’inhibiteur suicide.
Ils provoquent une destruction d’un groupement essentiel à l’activité donc l’enzyme devient
incapable de fixer ou transformer le substrat (ça n’existe pas dans le corps mais intérêt en
thérapeutique, exemple des AINS)
b) Inhibitions réversibles
Il y en a 3 types : compétitive , non compétitive et incompétitive. La fixation de l’inhibiteur
sur l’enzyme se fait par des liaisons faibles (formation et rupture faciles et rapides).
Inhibition compétitive
L’inhibiteur a une analogie structurale avec le substrat donc il se fixe au niveau du site actif à
la place du substrat

Il y a une levée de l’inhibition par excès de substrat : déplacement de l’équilibre entre EI et E.
est inchangé, augmente donc il y a diminution de l’affinité apparente pour S.
Inhibition non compétitive
Il n’y a pas d’analogie structurale avec le substrat donc la fixation de l’inhibiteur se fait sur
un site distinct du substrat (site actif). L’inhibiteur entraîne un changement de conformation
en se fixant à l’enzyme et diminue ainsi la vitesse de réaction. L’inhibiteur a la même affinité
sur forme libre E et forme complexée ES, indépendamment de S, une fixation simultanée de
I et S est possible. diminue, inchangé donc l’affinité pour S est inchangée.
Inhibiton incompétitive
Il y a une fixation sur forme ES (E déjà complexée au S), on a alors un équilibre E + S ES
déplacé en faveur de ES et l’affinité apparente de E pour S augmente donc diminue mais
le complexe ESI n’évolue pas donc diminue.
 6
7
6
7
1
/
7
100%