DC-SIGN et infections DC-SIGN and infections M

120
La Lettre de l’Infectiologue - Tome XIX - n° 4 - juillet-août 2004
M
I S E A U P O I N T
es découvertes récentes dans le domaine de l’immu-
n o l ogie permettent de mieux compre n d re les réactions
de défense mises en jeu par un hôte vis-à-vis d’un
agent infectieux. Pour éliminer les micro - o rganismes pat h o-
gènes (bactéries, virus, champignons et parasites), le système
i m mu n i t a i re utilise deux systèmes de défe n s e :l ’ i m mu n i t é
innée, dite naturelle,peu spécifique ou non adaptative, et l’im-
munité spécifique, dite adaptative. Ces deux systèmes ne fonc-
tionnent pas de façon indépendante et l’immunité naturelle a
un rôle d’information qui permet à la réponse spécifique d’être
a d aptée au type d’infe c t i o n : réponse cellulaire vis-à-vis des
ge rmes intra c e l l u l a i re s , réponse humorale vis-à-vis des ge rm e s
ex t r a c e l l u l a i res. Dans ce dialogue entre l’immunité innée et
l’immunité acquise,les cellules dendritiques (DC) ont un rôle
primordial, puisqu’elles ont la faculté de capturer puis de pré-
senter l’antigène aux cellules effectrices de la réponse immu-
nitaire (lymphocytes T et B). Récemment, un certain nombre
de travaux axés sur la biologie des DC ont permis de mieux
comprendre les modes d’interactions entre ces dernières et les
différentes classes d’agents infectieux, notamment à travers les
études réalisées sur un récepteur situé à la surface des DC ap p e l é
DC-SIGN (dendritic cell-specific ICAM 3-grabbing non-inte -
grin). Cet article est une mise au point sur le rôle de DC-SIGN
dans les interactions agents infectieux/cellules dendri t i q u e s .
Dans le système immunitaire, les DC ont un rôle essentiel de
cellules présentatrices d'antigène (CPA) (1, 2). Leur fonction
i m mu n o s t i m u l a t rice est désormais bien connu e : ces cellules
sont capables d’activer la réponse innée et la réponse adapta-
tive. Les DC tissulaires, présentes dans tous les tissus, captu-
rent et découpent l’antigène pour l’ap p r ê t e r, sous fo rme de pep-
t i d e s , en association avec les molécules de classes I et II du
c o m p l exe majeur d’histocompatibilité (CMH). Au cours de
cette succession d’événements, les DC migrent du site de cap-
t u re de l’antigène ve rs le cort ex pro fond des ganglions et acquiè-
rent ou augmentent l’ex p ression de molécules de costimu l a t i o n
(i.e. CD80, CD86) et de présentation (CMH de classe I et II),
p e r mettant de facto l’interaction avec des ly m p h o cytes T “ n a ï f s ”
et l’induction d’une réponse immu n i t a i re T spécifique ( 1 ) .
Outre leur capacité d’activation lymphocytaire, les DC contrô-
lent la différenciation des lymphocytes T CD4+ vers les voies
Th1 ou Th2 chez l'homme. Il existe différentes populations de
D C , chacune ayant ses pro p res précurs e u rs (pDC), se diff é-
DC-SIGN et infections
DC-SIGN and infections
J. F. Viallard*, F. Halary**, P. Blanco***, J. F. Moreau***, J.L. Pellegrin*
* Service de médecine interne et maladies infectieuses, hôpital Haut-Lévêque,
33604 Pessac.
** GRIV, équipe Avenir INSERM, université Bordeaux-2, 33076 Bordeaux.
*** CIRID, CNRS UMR 5164, université Bordeaux-2, 33076 Bordeaux.
RÉSUMÉ.
Dans le dialogue entre l’immunité innée et l’immunité acquise,les cellules dendritiques (DC) ont un rôle primordial, puisqu’elles
ont la faculté de capturer puis de présenter l’antigène aux cellules effectrices de la réponse immunitaire (lymphocytes T et B). Récemment, un
certain nombre de travaux axés sur la biologie des DC ont permis de mieux comprendre les modes d’interactions entre ces dernières et les
différentes classes d’agents infectieux, notamment à travers les études réalisées sur un récepteur situé à la surface des DC appelé DC-SIGN
(dendritic cell-specific ICAM 3-grabbing non-integrin). DC-SIGN semble jouer un rôle fondamental dans la capture et la transmission de
plusieurs agents infectieux, notamment le virus de l’immunodéficience humaine,le cytomégalovirus et Mycobacterium tuberculosis. Cette
lectine représente une cible de choix pour le développement de stratégies thérapeutiques visant à bloquer très précocement l'infection des DC
par les pathogènes capables d’interagir avec elle.
Mots-clés :
DC-SIGN - VIH - Cytomégalovirus - Mycobacterium tuberculosis.
ABSTRACT.
Dendritic cells (DC) have evolved to monitor the environment, detect pathogens and trigger T cell activation. Recent papers
focused on DC-SIGN (dendritic cell-specific ICAM 3-grabbing non-integrin), a C-type lectin which is is specifically (though not exclusively)
expressed on DCs, show that this new molecule mediates the capture and transfer of several infectious agents such as HIV, cytomegalovirus
and Mycobacterium tuberculosis. These findings could form the basis for novel therapeutic strategies aimed at blocking DCs infection by infec-
tious pathogens.
Keywords:
DC-SIGN - HIV - Cytomegalovirus - Mycobacterium tuberculosis.
L

La Lettre de l’Infectiologue - Tome XIX - n° 4 - juillet-août 2004
121
M
I S E A U P O I N T
renciant en DC immat u res puis mat u res. Les DC mat u res se
localisent dans la zone T des organes lymphoïdes secondaires
et ont pour fonction essentielle d'activer les ly m p h o cytes T naïfs
a fin d'induire une réponse immu n i t a i re spécifique ( 1 ) . C e t t e
réponse T peut être de type Th1, avec sécrétion d'interféron γ
( I F Nγ) ou d’interl e u k i n e 2 (IL-2) stimulant de manière pré-
pondérante l’immunité cellulaire, ou de type Th2, avec sécré-
tion d'interleukines (IL-4, I L - 5 , IL-10) stimulant l’immu n i t é
humorale (3).
Deux populations distinctes de DC humaines, nommées DC1
et DC2, contrôlent la différenciation des cellules T naïves en
ly m p h o cytes Th1 ou Th2. In vitro , les DC1, o b t e n ues par la
culture de monocytes en présence de GM-CSF (granulocyte-
macrophage colony-stimulating factor) et d’IL-4, puis activées
par le ligand du CD40 (CD40-L) (4), induisent la différencia-
tion de lymphocytes T naïfs CD4+CD45RA+ en lymphocytes
Th 1 , essentiellement par l’interm é d i a i r e d’une sécrétion
d’IL-12. Les DC2, générées in vitro à partir de cellules plas-
mocytoïdes CD4+CD3-CD11c- (pDC2) cultivées six jours en
présence d’IL-3, puis activées par le CD40-L (5), induisent une
différenciation de type Th2. Dans le modèle actuel de la diffé-
re n c i ation Th 1 / Th 2 , l’IL-12 est le principal facteur de diff é-
renciation Th1, tandis que l’IL-4 est essentielle à l’induction
d’une réponse Th2 (3). Or les DC2 ne sécrètent pas d’IL-12.
Ainsi, l’orientation Th1 ou Th2 de la cellule T naïve serait en
p a rtie déterminée par le type de DC (DC1 ou DC2) présent dans
un microenvironnement donné.
Il est bien établi que les DC in vivo résident dans les tissus
dans un état quiescent et nécessitent des signaux ex t é ri e u rs ,
q u a l i fiés de “signaux de dange r ” , pour acquérir un état effe c-
teur dans lequel elles peuvent alors induire une réponse cellu-
l a i re T. Ce processus d’activation des DC semble être dire c t e-
ment régulé par la perc eption des pat h ogènes par les cellules
elles-mêmes. Cette perc eption est largement dépendante de
r é c e p t e u r s appelés Pat t e r n Recognition Recep t o rs( P R R ) ,
c ap abl es de re c o n n a î t re des motifs moléculaires conserv é s .
Ces PRR sont représentés par deux grandes fa m i l l e s : les récep-
t e u rs t o l l - l i ke (TLR) et les lectines de type C (C car actives en
présence d’ions calcium). Selon le type de tissu dans lequel
elles sont localisées et leur degré de diff é r e n c i at i o n , les DC
sont spécialisées dans une réponse à un type d’agent micro-
bien bien spécifique simplement parce qu’elles ex p riment dif-
f é rents types de PRR, eux-mêmes très spécifiques de ce pat h o-
g è n e. Si les TLR reconnaissent plutôt des lipopro t é i n e s , d e s
l i p o p o ly s a c ch a rides (LPS) ou de l’ADN bactérien déméthy l é
(CpG DNA) ( 6 ) , les lectines de type C reconnaissent des rési-
dus sucrés, p o rtés uniquement par certaines classes de pat h o -
gènes ou certaines gly c o p r otéines du soi ( 7 ) . A l o r s que les
T L R , en interagissant avec leurs liga n d s , ont la capacité de
t ra n s m e t t re un signal d’alerte à la cellule, par toute une cas-
cade d’événements moléculaires concrétisés in fine par la
sécrétion de cytokines ou l’ex p res sion de molécules de costi-
mu l at i o n , les lectines de type C ont surtout pour fonction de
c ap t u r er et d’internaliser les antigènes re c o n nu s , avant de les
présenter classiquement en surface ( 8 ).
À ce jour,peu de lectines de type C ont été clairement impli-
quées dans la perc eption d’agents pat h ogènes identifi é s .
Récemment, l’une d’entre elles, connue sous le nom de DC-
SIGN (dendritic cell-specific ICAM 3-grabbing non-integrin)
ou CD209, a été très étudiée en raison, notamment, de son affi-
nité pour la gly c o p rotéine gp120 de l’enveloppe du VIH ( 9 , 1 0 ) .
DC-SIGN est une gly c o p rotéine membra n a i re de 44 kDa, ex p ri-
mée sur les DC immatures et matures. C’est un récepteur natu-
rel de ICAM-2 et ICAM-3 (intercellular adhesion molecule-
2 / 3 ) , mais elle ne fait cependant pas partie de la famille des
intégrines. Contrairement aux autres lectines de type C, telles
que DEC205 ou le récepteur au mannose (MR), qui possèdent
respectivement 8 et 10 domaines de reconnaissance des carbo-
hydrates (CRD), DC-SIGN n’en contient qu’un seul. L’inter-
action entre DC-SIGN et ICAM-2 ex p rimé sur les cellules endo-
théliales conditionne le roulement, l’attachement et finalement
l ’ ex t rava s a tion des DC immat u res du vaisseau ve rs le foye r
i n fl a m m at o i re. L’ i n t e r action entre DC-SIGN et ICAM-3 (ex p ri-
mée de façon prépondérante par les ly m p h o cytes T au rep o s
dans les ganglions lymphatiques) permet aux DC matures de
renforcer la zone de contact étroit, appelée “synapse immuno-
logique”, nécessaire au déclenchement de l’activation des lym-
phocytes T naïfs.
DC-SIGN ET VIH
Dans le contexte infectieux, l’intérêt porté à la molécule DC-
SIGN est venu des travaux menés par Geijtenbeek et al. sur le
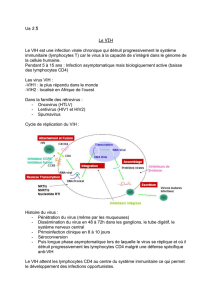
VIH ( 9 , 1 0 ) . Comme les monocy t e s / m a c ro p h age s , les DC ex p r i-
ment le récepteur CD4 et le corécepteur CCR5, à des niveaux
toutefois plus faibles que les lymphocytes T CD4+, ces deux
molécules formant un complexe qui permet la fusion du VIH
avec la membrane cellulaire et son entrée dans la cellule. Il
semble que les DC des muqueuses jouent un rôle de porte d’en-
trée du virus et de cheval de Troie du VIH, puisqu’elles sont
c ap ables de le tra n s p o rter jusqu’aux zones T des ganglions dra i-
nants (11).
L’infection productive des DC en l’absence de lymphocytes T
a été mise en évidence in vivo (12, 13). La réplication du virus
n’est possible que dans les DC immatures, mais elles peuvent
le transmettre à des lymphocytes T et, en les stimulant, créer
les conditions d’une production virale explosive (12). La liai-
son de la gp120 à des DC immatures est inhibée par des anti-
c o rps anti-DC-SIGN, a l o rs que des anticorps anti-CD4 sont
inefficaces. DC-SIGN est nécessaire à l’infection des lympho-
cytes T dans des cocultures de DC et de ly m p h o cyt es T.
DC-SIGN ne se substitue pas au complexe CD4/CCR5 pour
l’entrée du virus dans la cellule,et à lui seul ne la permet pas,
du moins dans des cellules perm i s s ives pour la réplication. Dans
des ex p é riences in vitro , il a été montré que les DC peuve n t
c apter les particules virales via le récepteur DC-SIGN et les
stocker plusieurs jours dans un compartiment cellulaire encore
mal défini, sans qu’elles perdent leur pouvoir infectieux (14).
DC-SIGN reconnaît la gp120 de VIH-1 et internalise le virus,
mais ne le dégrade pas pendant au moins quatre jours, si bien
que ce dernier garde son caractère infectieux, participant ainsi

122
La Lettre de l’Infectiologue - Tome XIX - n° 4 - juillet-août 2004
M
I S E A U P O I N T
à l’échappement immunitaire. Cela laisse le temps à la DC de
le tra n s p o r ter ve r s les régions T des organes lymphoïdes. Quand
les DC rencontrent une cellule T, VIH-1 retourne à la surface
c e l l u l a i re et DC-SIGN le transmet aux ly m p h o c ytes T, a u g-
mentant ainsi l’infection de ces cellules. De plus, dans les DC
infectées, la protéine Nef de VIH-1 perturbe le parcours intra-
cellulaire de DC-SIGN,ce qui entraîne une forte augmentation
des taux de DC-SIGN à la surface des DC, avec comme consé-
quence une amplification de l’adhésion aux cellules T et de la
transmission du VIH (15). Enfin, DC-SIGN, après avoir per-
mis la cap t u re du virus par les DC, facilite l’infection “en tra n s ”
de cellules permissives à la réplication.
DC-SIGN permet la cap t u re du virus par les DC subépithéliales
des muqueuses et facilite son transport vers les régions T des
organes lymphoïdes secondaires où a lieu l’infection des lym-
p h o cytes T CD4+. Il est important de noter ici que les DC ex p r i-
mant DC-SIGN sont entre autres localisées in vivo près de la
lamina propria des muqueuses du rectum, de l’utérus et du col
u t é r in. Elles peuvent cap t u rer les virions ayant un tropisme pour
les corécepteurs CCR5 et CXCR4 et infecter secondairement
des cellules permissives. Or,in vivo, il existe une transmission
préférentielle des virus ayant un tropisme pour CCR5. L’inter-
action avec un autre type de DC n’exprimant à leur surface que
C C R 5 , et pas DC-SIGN, comme les cellules de Lange r h a n s
(DC des épithéliums malpighiens) ou les monocy t e s / m a c r o-
phages, pourrait expliquer la restriction de la transmission aux
souches R5.
DC-SIGN ET CYTOMÉGALOVIRUS (CMV)
L’ i n fection par le CMV (ou HHV-5) est répandue dans le monde
entier (50-90 % de prévalence). Ce virus peut être mis en évi-
dence dans le sperme, l’urine, la salive, les fèces, les sécrétions
situées au niveau du col de l’utérus (surtout pendant la gros-
sesse), et dans le lait maternel. Le CMV peut devenir un pro-
blème dans trois circonstances principales. Chez la fe m m e
e n c e i n t e, la transmission fœtomat e rn e l l e , avec un cas pour
1 0 0 naissances et des séquelles dans env i ron un cas pour
10 0 0 n a i s s a n c e s , fait du CMV une des pre m i è res causes d’em-
bryo-fœthopathie infectieuse. Chez les malades infectés par le
VIH et au cours d’autres immu n o s u p p re s s i o n s , des at t e i n t e s
viscérales spécifiques (rétiniennes, digestives...) sont décrites.
Chez les greffés, le CMV pourrait être impliqué dans le rejet
ch ron ique de gre ffons solides (16). M a l g ré la somme de
connaissances accumulée au cours des trente dernières années,
depuis sa mise en évidence,le CMV reste mystérieux et fidèle
à sa devise, qu’il partage avec tous les autres herpèsvirus : pour
vivre heureux, vivons cachés !
Comme les virus de l’immu n o d é fici ence humaine, d ’ E b o l a
(17), de l’hépatite C (18) et de la dengue (19), le CMV humain
(HCMV) a également la capacité d’interagir avec les DC via
D C - S I G N,et au-delà, de participer activement aux événements
précoces de la physiopathologie de l’infection à CMV (20). À
l’instar du VIH, le blocage in vitro de DC-SIGN,par des anti-
corps neutralisants, inhibe non seulement la capture du CMV
par les DC, mais aussi la transmission du virus vers d’autres
types cellulaires permissifs (phénomène de “trans-infection”).
La neutra l i s ation de DC-SIGN empêche en outre l’infe c t i o n
des DC par des isolats cliniques de CMV. Des expériences uti-
lisant une lignée cellulaire DC-SIGN positive et fa i blement sus-
c ep t i ble au CMV (i.e. le virus infecte la cellule, mais ne se mu l-
tiplie pas – pas de cy cle ly t i q u e ) , comparée à la lignée pare n t a l e
– DC-SIGN négat ive – ont montré que l’ex p r ession de DC-
SIGN suffisait, dans ce cas, pour rendre la cellule complète-
ment permissive au virus (i.e. le virus rentre et se multiplie –
lyse de la cellule). Les mécanismes exacts de ce phénomène
(“cis-infection”) sont encore inconnus, mais ces données sug-
gèrent déjà que DC-SIGN a un rôle facilitateur de l’infection
CMV. Les résultats que nous avons obtenus apportent un éclai-
rage nouveau sur les mécanismes moléculaires impliqués dans
l’infection des DC par le CMV, surtout lorsque l’on sait que le
CMV a des pro p riétés immu n o s u p p r e s s ives fo rmellement docu-
mentées (21). Brièvement, l’infection d’un hôte par le CMV
est nécessaire au déclenchement d’un programme codé par le
génome viral visant à diminuer les capacités de défense de
l ’ h ô t e. Des travaux récents ont démontrés que les DC pouva i e n t
ê t re infectées in vitro par le CMV, à condition d’utiliser une
souche primaire, non adaptée à la culture in vitro (i.e. isolat cli-
nique) et des DC immatures. Dans ces conditions expérimen-
tales stri c t e s , les auteurs de ces trava u x , e f fectués chez l’homme,
ont démontré que des DC infectées par le CMV voyaient leurs
fo n c t i o n s , p rincipalement de stimu l ation des ly m p h o cytes T,
fo rtement affectées ( 2 2 ) . Les mécanismes conduisant à cette
perte de la fonctionnalité des DC par l’infection CMV reposent
p rincipalement sur la diminution de l’ex p ression des molécules
costimulatrices (i.e. CD80, CD86) et des molécules du CMH
de classe I (rétention dans le réticulum endoplasmique par la
glycoprotéine virale gpUS3, translocation dans le cytoplasme
et dégradation par gpUS2/11 et, enfin, inhibition de TAP et du
chargement des peptides par gpUS6) et II (inactivation virale
du tra n s - a c t ivateur des molécules de classe II, C I I TA) ( 2 3 ) .
DC-SIGN ET INFECTIONS À MYCOBACTÉRIES
Très récemment, deux groupes indépendants ont montré que
les mycobactéries (en particulier Mycobacterium tuberculosis)
utilisaient également DC-SIGN par l’intermédiaire d’un motif
o l i g o-mannosylé de l’extrémité d’un lipog lycane ab o n d a m m e n t
présent dans la paroi cellulaire des mycobactéries virulentes à
croissance lente,le Man-LAM (mannose-capped lipoarabino -
mannan) (24, 25). Man-LAM est reconnu par DC-SIGN, mais
également par d’autres récepteurs tels que CD11b ou CD11c.
Le CD11 fait partie du récepteur au complément de type 3
(CR3) qui fixe les mycobactéries soit directement par un site
de liaison au mannane,soit indirectement en fixant des com-
posants du complément activé présents sur la surface bacté-
rienne. Néanmoins, grâce à des études menées avec des anti-
corps bloquants, il semble que la liaison des mycobactéries aux
DC ne se fasse que par DC-SIGN,et non avec les autres récep-
t e u r s. Man-LAM est un motif absent ou modifié dans les
souches saprophytes ou d’autres souches de mycobactéries à
c roissance lente (comme M y c o b a c t e r ium av i u m). DC-SIGN

La Lettre de l’Infectiologue - Tome XIX - n° 4 - juillet-août 2004
123
M
I S E A U P O I N T
permettrait ainsi au système immunitaire de faire la différence
entre des mycobactéries virulentes et des mycobactéries sapro-
phytes ou opportunistes.
Les DC ont été localisées dans la sous-muqueuse et les espaces
i n t e rstitiels du tractus re s p i r at o i re. A i n s i , elles pourraient dire c-
tement capter M. tuberc u l o s i s et tra n s p o r ter cet agent pat h o-
gène du site pri m a i re de l’infection jusqu’au ganglion ly m-
p h atique qui le draine (complexe de Ghon). Comme pour le
V I H , on peut supposer que les my c o b a c t é ries du complexe
tuberculosis utilisent les DC comme un cheval de Troie, afin
de se disséminer dans l’organisme et d’atteindre les ganglions
ly m p h atiques. À ce nive a u , les DCs présentent les antigènes
my c o b a c t é riens aux ly m p h o c ytes T qui, en réponse, vo n t
induire une réponse protectrice Th1 via l’IFNγavec activation
des macro p h ages et des ly m p h o cytes cy t o t oxiques (qui libère n t
une combinaison de perfo rine et de gra nu ly s i n e / g ra n z y m e ) .
Tailleux et al. ont montré que les DC du complexe de Ghon
exprimaient DC-SIGN et que les DC positives pour DC-SIGN
p rovenant de patients tuberculeux étaient porteuses des anti-
gènes de M. tuberculosis, ce qui fait le lien entre l’observation
in vitro et la situation in vivo (25).
On sait que les my c o b a c t é ries sont de puissants inducteurs
de la voie Th 1 , et plusieurs de leurs composants stimulent
l’expression des molécules de costimulation et la production
d’IL-12 dans les DC via TLR2 et TLR4. Inve rs e m e n t , L A M
stimule la production d’IL-10, cytokine immunosuppressive et
anti-inflammatoire, et, en même temps, diminue la maturation
des DC induite par le LPS. En conséquence,on peut donc ima-
giner que la voie pro-inflammatoire, d’un côté, résulterait d’un
mécanisme destiné à limiter les séquelles pathologiques. En ce
s e n s , DC-SIGN (et peut-être d’autres récep t e u rs de LAM) pour-
rait contre c a r rer l’activation médiée par les TLR. Les my c o -
bactéries, cependant, pourraient exploiter cette voie suppres-
s ive en faisant basculer l’équilibre précaire des réponses
pro-inflammatoires protectrices/réponses anti-inflammatoires
suppressives en faveur de ces dernières, et pour leur plus grand
profit.
CONCLUSION
Le rôle fondamental de DC-SIGN dans la capture et la trans-
mission (cis- et/ou trans) du CMV par les DC fait de cette lec-
tine une cible de choix pour le développement de strat é g ies thé-
rapeutiques visant à bloquer très précocement l’infection des
CD par le CMV, ainsi que par tous les autres pat h og è n e s
capables d’interagir avec DC-SIGN.
R
É F É R E N C E S B I B L I O G R A P H I Q U E S
1.
Banchereau J,Steinman RM. Dendritic cells and the control of immunity.
Nature 1998;392:245-52.
2.
Clark GJ,Angel N,Kato M et al. The role of dendritic cells in the innate immu-
ne system. Microbes Infect 2000;2:257-72.
3.
O’Garra A. Cytokines induce the development of functionally heterogeneous T
helper cell subsets. Immunity 1998;8:275-83.
4.
Sallusto F, L a n z ave c chia A. Efficient pre s e n t ation of soluble antigen by culture d
human dendritic cells is maintained by granulocyte/macrophage colony-stimula-
ting factor plus interleukin 4 and downregulated by tumor necrosis factor alpha.
J Exp Med 1994;179:1109-18.
5.
Grouard G,Rissoan MC, Filgueira L, Durand I, Banchereau J,Liu YJ. The
enigmatic plasmacytoid T cells develop into dendritic cells with IL-3 and CD40-
ligand. J Exp Med 1997;185:1101-11.
6.
Underhill DM, Ozinsky A. Toll-like receptors: key mediators of microbe detec-
tion. Curr Opin Immunol 2002;14:103-10.
7.
D ri ckamer K. C-type lectin-like domains. Curr Opin Struct Biol 1999;9: 5 8 5 -
9 0 .
8.
Figdor CG, van Kooyk Y, Adema GJ. C-type lectin receptors on dendritic cells
and Langerhans cells. Nat Rev Immunol 2002;2:77-84.
9.
Geijtenbeek TB, Torensma R, van Vliet SJ et al. Identification of DC-SIGN,
a novel dendritic cell-specific ICAM-3 receptor that supports primary immune
responses. Cell 2000;100:575-85.
10.
Geijtenbeek TB, Kwon DS, Torensma R et al. DC-SIGN,a dendritic cell-spe-
cific HIV-1-binding protein that enhances trans-infection of T cells. Cell 2000;
100:587-97.
11.
Spira AI, Marx PA, Patterson BK et al. Cellular targets of infection and route
of viral dissemination after an intravaginal inoculation of simian immunodefi-
ciency virus into rhesus macaques. J Exp Med 1996;183:215-25.
12.
Hosmalin A. Subornation des lymphocytes T naïfs par un nouveau récepteur
des cellules dendritiques. Médecine/Sciences 2000;16:1121-3.
13.
Steinman RM. DC-SIGN : a guide to some mysteries of dendritic cells. Cell
2000;100:491-4.
14.
Kwon DS, Gregorio G,Bitton N, Hendrickson WA, Littman DR. DC-SIGN-
mediated internalization of HIV is required for trans-enhancement of T cell infec-
tion. Immunity 2002;16:135-44.
15.
Sol-Foulon N,Moris A, Nobile C et al. HIV-1 Nef-induced upregulation of
DC-SIGN in dendritic cells promotes lymphocyte clustering and viral spread.
Immunity 2002;16:145-55.
16.
Alain S, Mazeron MC. Cytomégalovirus. Traité de virologie médicale. Édi-
tions ESTEM 2003;195-211.
17.
Alvarez CP, Lasala F, Carrillo J, Muniz O, Corbi AL, Delgado R. C-type lec-
tins DC-SIGN and L-SIGN mediate cellular entry by Ebola virus in cis and in
trans. J Virol 2002;76:6841-4.
18.
Tassaneetrithep B, Burgess TH, Granelli-Piperno A et al. DC-SIGN (CD209)
mediates dengue virus infection of human dendritic cells. J Exp Med 2003;
197:823-9.
19.
Gardner JP,Durso RJ,Arrigale RR et al. L-SIGN (CD 209L) is a liver-spe-
cific capture receptor for hepatitis C virus. Proc Natl Acad Sci USA 2003;100:
4498-503.
20.
Halary F, Amara A, Lortat-Jacob H et al. Human cytomegalovirus binding to
DC-SIGN is required for dendritic cell infection and target cell trans-infection.
Immunity 2002;17:653-64.
21.
Naniche D. Generalized immunosuppression : individual viruses, intertwined
targets. Virology 2000;275:227-32.
22.
Raftery MJ,Schwab M, Eibert SM, Samstag Y, Walczak H, Schonrich G.
Targeting the function of mature dendritic cells by human cytomegalovirus: a
multilayered viral defense strategy. Immunity 2001;15:997-1009.
23.
Miller DM, Rahill BM, Boss JM et al. Human cytomegalovirus inhibits major
histocompatibility complex class II expression by disruption of the Jak/Stat
pathway. J Exp Med 1998;187:675-83.
24 .
Geijtenbeek T B , van Vliet SJ, Koppel EA et al. Mycobacteria tar-
get DC-SIGN to suppress dendritic cell function. J Exp Med 2003;197:
7 - 1 7 .
25.
Tailleux L, Schwartz O, Herrmann JL et al. DC-SIGN is the major
Mycobacterium tuberculosis receptor on human dendritic cells. J Exp Med 2003;
197:121-7.

?
?
124
La Lettre de l’Infectiologue - Tome XIX - n° 4 - juillet-août 2004
M
I S E A U P O I N T
F
ormation
M
édicale
C
ontinue
M
C
?
I.
Parmi ces virus, quel(s) est (sont) celui (ceux) qui
est (sont) capté(s) par DC-SIGN ?
a. le virus d’Epstein-Barr
b. le Cytomégalovirus
c. l’herpèsvirus 1
d. l’herpèsvirus 8
e. le virus VIH
f. le virus de la dengue
a. les lipopolysaccharides
b. des résidus sucrés
c. l’ADN bactérien déméthylé
d. des lipoprotéines
a. les DC expriment le récepteur CD4 et le corécepteur CCR5,
permettant ainsi l’entrée du VIH dans ces cellules
b. la réplication du VIH n’est possible que dans les DC matu r e s
c. les DC des muqueuses sont capables de transporter le VIH
jusqu’aux zones T des ganglions drainants
d. les DC exprimant DC-SIGN sont surtout localisées in vivo
près de la lamina propria des muqueuses du rectum, de
l’utérus et du col utérin
II.
Quel type de structure reconnaissent les lectines
de type C ?
I I I .
Concernant le VIH, quelle(s) proposition(s) est
(sont) vraie(s) ?
w w w. v i h . g s k . f r
1
/
5
100%