Grosset_Andree-Anne_2009_memoire


Université de Montréal
Caractérisation du potentiel métastatique des carcinomes mammaires
de phénotype basal
par
Andrée-Anne Grosset
Département de pathologie et biologie cellulaire
Faculté de Médecine
Mémoire présenté à la Faculté des études supérieures
en vue de l’obtention du grade de M.Sc.
en Pathologie et biologie cellulaire
option Biologie du cancer
Août, 2009
©, Andrée-Anne Grosset, 2009

Université de Montréal
Faculté des études supérieures
Ce mémoire intitulé :
Caractérisation du potentiel métastatique des carcinomes mammaires
de phénotype basal
présenté par :
Andrée-Anne Grosset
a été évalué par un jury composé des personnes suivantes :
Roger Lippé
président-rapporteur
Louis Gaboury
directeur de recherche
Yves St-Pierre
codirecteur
Nicole Leclerc
membre du jury

i
RÉSUMÉ ET MOTS CLÉS
Au Canada, on estime qu'une femme sur neuf développera un cancer du sein et qu'une
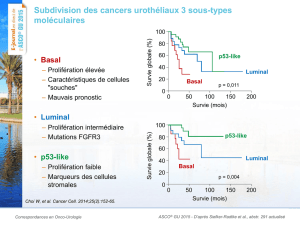
femme sur vingt-huit en mourra. Les carcinomes mammaires de phénotype basal qui
comprennent environ 15 à 25% des cancers envahissants du sein sont des tumeurs malignes
ayant un très mauvais pronostic. Dans ce type tumoral, les métastases sont fréquentes et les
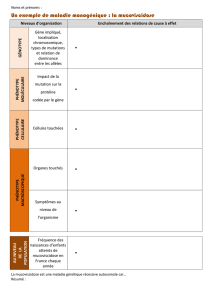
décès nombreux. La formation des métastases est un processus complexe qui comprend
plusieurs étapes; chacune peut être étudiée par des marqueurs spécifiques. Notre hypothèse
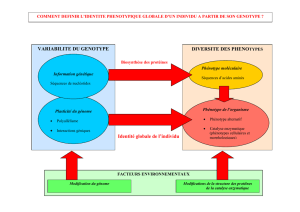
de recherche est que le carcinome mammaire de phénotype basal possède des
caractéristiques spécifiques qui permettent d'expliquer son potentiel métastatique élevé.
Six micromatrices tissulaires comprenant un total de 213 tumeurs mammaires ont été
confectionnées. L'expression des marqueurs usuels de la métastase et celle de nouveaux
marqueurs a été étudiée par des techniques d'immunohistochimie.
L'étude comparative de l’expression des marqueurs du potentiel métastatique par les
carcinomes mammaires de phénotype basal indique que les protéines Ki-67, EGFR, CD276
et galectine-7 sont étroitement reliées à ce type de cancer. De plus, l'expression du
marqueur GATA-3, un marqueur anti-métastatique, fait complètement défaut. Nos résultats
confirment que le cancer du sein de phénotype basal possède un plus grand potentiel
métastatique que les autres sous-types génétiques et suggèrent que la galectine-7 puisse être
impliquée.
Mots clés : cancer du sein, immunohistochimie, métastases, micromatrice tissulaire.

ii
ABSTRACT AND KEYWORDS
In Canada, we estimate that one woman in nine will develop a breast cancer, and one
woman in twenty-eight will succumb of her disease. The basal-like breast cancer which
represents 15 to 25% of invasive breast cancer, is associated with a poor prognosis.
Metastases are the main cause of mortality for patients affected from cancer. Many steps
are involved in the metastatic cascade. Each step can be studied using specific markers. Our
research hypothesis is that basal-like breast cancer have distinct charactheristics which can
explain their higher metastatic potential and poor overall survival.
Six tissue microarrays comprising a total of 213 breast tumors were assembled. The relative
expression of conventional and putative metastasis related markers was studied using
immunohistochemistry.
When the expression of metastatic potential markers was evaluated in breast carcinomas,
basal-like breast cancer was found to express higher levels of Ki-67, EGFR, CD276 and
galectin-7. Furthermore, GATA-3, an antimetastatic protein, was found to be completely
absent in basal-cell type breast cancers. Our results confirm that the basal-like breast cancer
has a highest metastatic potential than the other molecular subtypes and suggest that
galectin-7 may play a role.
Keywords : basal-like subtype, breast cancer, immunohistochemistry, metastasis, tissue
microarray.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
1
/
151
100%