Brèves… Brèv es… Le PYY

289
Métabolismes Hormones Diabètes et Nutrition (VII), n° 6, novembre/décembre 2003
Brèves…
Brèves…
Le PYY3-36 : un avenir dans
le traitement de l’obésité ?
La régulation de l’homéostasie éner-
gétique met en jeu des circuits neuro-
endocriniens complexes. Le cerveau
est renseigné en permanence sur l’état
des réserves et des besoins énergé-
tiques de l’organisme par des hor-
mones circulantes, qui peuvent notam-
ment être classées en deux grandes
catégories. La première d’entre elles
inclut les hormones qui agissent à
court terme et gouvernent directe-
ment le rythme et la durée des repas.
La cholécystokinine et la ghréline
entrent dans cette catégorie en tant
que signaux de satiété et de stimu-
lateur d’appétit, respectivement. Le
deuxième groupe comprend les hor-
mones agissant à plus long terme et
assurant la stabilité du poids corporel,
comme la leptine et l’insuline, dont
les taux sont directement corrélés à la
masse de tissu adipeux. Lorsque les
réserves graisseuses de l’organisme
diminuent, la baisse des taux circu-
lants de leptine et d’insuline est per-
çue par le cerveau et traduite en une
augmentation de l’appétit et du méta-
bolisme jusqu’à la récupération du
poids initial.
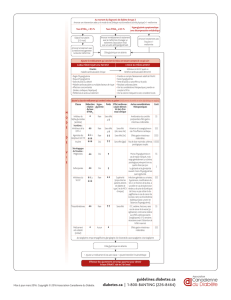
La cible centrale principale de ces fac-
teurs circulants est l’hypothalamus,
qui joue un rôle clé dans la régulation
de la prise alimentaire. Ce contrôle est
le résultat d’interactions complexes
entre deux réseaux neuro-peptider-
giques localisés dans le noyau arqué et
jouant le rôle d’accélérateur d’appé-
tit pour l’un, de frein pour l’autre.
Le système anorexigène synthétise
l’a-melanocyte-stimulating hormone
(a-MSH). Le système orexigène
coexprime le neuropeptide Y (NPY)
et l’agouti-related peptide (AgRP).
Un tiers des neurones à NPY/AgRP
produit également un autre facteur
orexigène, le GABA. Les neurones
à NPY/AgRP, activés par la perte de
poids, stimulent la prise de nourri-
ture de différentes manières : le NPY
libéré active les récepteurs Y1 au
niveau du noyau paraventriculaire.
Par ailleurs, le NPY exerce un contrôle
négatif sur les neurones anorexigènes
à a-MSH en inhibant l’expression du
gène codant le précurseur de l’a-MSH.
Le peptide AgRP se comporte quant à
lui comme un antagoniste des récep-
teurs de l’a-MSH de type centraux,
les MC4R. Ainsi, à la suite d’un jeûne
prolongé, les neurones à NPY sont
activés tandis que ceux à a-MSH sont
inhibés, ce qui entraîne une stimula-
tion de l’appétit et une récupération
des réserves énergétiques.
La clé des découvertes récentes dans
le contrôle de l’homéostasie énergé-
tique est la mise en évidence de récep-
teurs présynaptiques inhibiteurs Y2
(Y2R) sur les neurones NPY/AgRP.
Or, un agoniste puissant de ce récep-
teur est le PYY3-36, un autre membre
de la famille du NPY, qui partage
70 % d’identité avec le NPY. Le
PYY3-36 est libéré par les cellules
endocrines tapissant l’intestin grêle et
le côlon et sa concentration plasma-
tique est proportionnelle à la quan-
tité de calories ingérées. Sur la base
de ces observations, Batterham et al.
avaient supposé, lors d’une première
étude, que l’augmentation postpran-
diale des taux plasmatiques de PYY
pouvait inhiber la prise alimentaire
via l’activation des récepteurs Y2R
au niveau du noyau arqué. De fait, ils
ont montré que l’injection périphé-
rique de PYY inhibe la prise de nour-
riture et réduit le poids corporel chez
les rongeurs après une semaine de trai-
tement chronique (1). Chez l’homme,
l’administration de PYY3-36, à une
dose mimant les concentrations plas-
matiques du peptide en période post-
prandiale entraîne une réduction de
l’appétit et de la prise de nourriture
de 33 % sur une période de 12 heures.
En agissant à long terme, le PYY3-36
se distingue ainsi des autres hormones
de satiété du tractus gastro-intestinal,
qui agissent rapidement, comme la
cholécystokinine. Ils montrent, par
ailleurs, que le PYY3-36 influence
directement les circuits neuronaux du
noyau arqué. En se liant aux récep-
teurs Y2, le PYY3-36 hyperpolarise
les neurones à NPY/AgRP et inhibe la
libération de NPY. Le PYY3-36 exerce
également de façon indirecte une
activation de la libération d’a-MSH
par une levée du tonus inhibiteur
exercé par le GABA sur les neu-
rones à a-MSH. L’effet du PYY3-36
est mimé par le Y2A, un agoniste du
récepteur Y2, et aboli chez les souris
dont le gène Y2R est invalidé, deux
résultats qui démontrent bien l’im-
plication du récepteur Y2. Ainsi, le
NPY et le PYY3-36, deux peptides de
la même famille, exercent par l’inter-
médiaire de deux récepteurs diffé-
rents, Y1R et Y2R, des effets oppo-
sés sur la régulation de la prise
alimentaire.
Dans un contexte où l’obésité est
une cause majeure de maladies et de
décès à travers le monde (on compte
actuellement environ 280 000 décès
liés à l’obésité par an aux États-
Unis), l’effet anorexigène du pep-
tide PYY3-36 chez l’homme s’avère
extrêmement intéressant, compte tenu
de la résistance à la leptine observée
chez la plupart des sujets obèses.
L’équipe londonienne a donc naturel-
lement poursuivi ses travaux en tes-
tant l’efficacité du peptide PYY3-36
sur des sujets obèses (2). Les effets
du PYY3-36 sur l’appétit et la prise
alimentaire ont été comparés sur des
cohortes composées de sujets obèses
(6 hommes, 6 femmes) et de volon-
taires sains. Ils ont constaté une dimi-
nution de 30 % de la quantité de
calories ingérées lors d’un repas, à
la fois chez les témoins et chez les
sujets obèses. La diminution de la
prise de nourriture persiste 12 heures
après l’administration du peptide.
Ces résultats démontrent donc que
l’obésité n’est pas associée à une
résistance au PYY3-36. Batterham et
al. ont ensuite expérimenté l’hypo-
thèse selon laquelle une déficience
de production de PYY3-36 pouvait
contribuer à la physiopathologie
de l’obésité. De fait, les taux plas-
matiques de PYY3-36 endogène sont
plus faibles chez les sujets obèses,
comparativement aux sujets nor-
maux, à jeun ou après un repas,
bien
que la quantité de calories ingérée
soit plus importante. En outre, ils
montrent que les taux plasmatiques
de PYY3-36 sont inversement cor-
rélés à l’index de masse corporelle

Brèves…
Brèves…
290
Métabolismes Hormones Diabètes et Nutrition (VII), n° 6, novembre/décembre 2003
chez les sujets à jeun. L’administra-
tion de PYY3-36 diminue les taux
plasmatiques de ghréline dans les
deux groupes étudiés, une action qui
pourrait contribuer à accroître l’effet
de satiété du PYY3-36. Ces résultats
suggèrent ainsi qu’une déficience
en PYY3-36 circulant soit impliquée
dans la physiopathologie de l’obésité.
Il reste maintenant à déterminer si
elle en constitue la cause ou la consé-
quence. Les travaux de cette équipe
ouvrent donc des perspectives nou-
velles dans le traitement de l’obésité,
soit par administration de PYY3-36
exogène, soit en stimulant la libéra-
tion du peptide endogène.
I. Lihrmann,
INSERM U413,
université de Rouen.
La mémoire métabolique
du rein chez le diabétique
de type 1
L’étude DCCT (Diabetes Control
and Complications Trial) a démontré,
chez les diabétiques de type 1, les
bénéfices d’un traitement optimisé
versus conventionnel sur le contrôle
de l’hémoglobine glyquée et la
réduction des complications micro-
vasculaires, notamment rénales. Au
terme de cette étude d’intervention
de 6,5 années, l’observation de la
cohorte a été poursuivie pendant
8 années dans le cadre de l’étude
EDIC (Epidemiology of Diabetes
Interventions and Complications) (1).
Pendant cette étude d’observation,
les sujets du groupe “contrôle opti-
mal” de DCCT ont été encouragés à
poursuivre le même traitement et les
sujets du groupe “traitement conven-
tionnel”, à débuter un traitement
optimalisé du fait des résultats du
DCCT. Le suivi du diabète et les choix
thérapeutiques n’étaient cependant
plus contrôlés par les investigateurs,
mais confiés aux praticiens habituels
des patients. À la fin de l’étude DCCT,
donc au début de l’étude EDIC,
l’HbA1c était à 7,2 % pour le groupe
“optimal” versus 9,1 % pour le
groupe “conventionnel” (p < 0,001) ;
cette différence a disparu dès la pre-
mière année de l’étude EDIC, avec
une élévation de l’HbA1c pour le
groupe “optimal” à 8 % et une dimi-
nution pour le groupe “convention-
nel” à 8,2 %. Malgré ce nivellement
des niveaux d’HbA1c entre les deux
groupes, le bénéfice, en termes de
diminution de l’incidence et de la
progression de la néphropathie dia-
bétique, a persisté au cours de l’étude
EDIC. En effet, pendant les 8 années
de cette étude, une micro-albuminurie
et une albuminurie ne sont survenues
respectivement que chez 6,8 % et
1,4 % des patients du groupe “opti-
mal”, versus 15,8 % et 9,4 % pour
le groupe “conventionnel”. Ainsi, le
groupe ayant bénéficié du traite-
ment optimisé dans l’étude DCCT
conserve une protection rénale 8 ans
après la fin de cette étude d’inter-
vention, et ce malgré une quasi-
égalisation de l’HbA1C des groupes
“traitement optimisé” et “traitement
conventionnel”. Ce bénéfice soulève
donc à nouveau l’hypothèse de la
“mémoire métabolique”, concept
déjà évoqué pour la rétinopathie
diabétique : en effet, le ralentisse-
ment de la progression de la rétino-
pathie diabétique, obtenu grâce à
un contrôle glycémique optimisé
versus conventionnel, persiste plu-
sieurs années après la dégradation
du contrôle glycémique par relâ-
chement du traitement optimal (2).
Cet effet rémanent du bon contrôle
glycémique sur la progression de la
maladie microvasculaire diabétique
pourrait être en partie expliqué par
les AGE (Advanced Glycation End
products). En effet, l’accumulation
de ces AGE dans les tissus cibles est
corrélée au contrôle glycémique et
semble prendre une part importante
dans la physiopathogénie des lésions
microvasculaires.
Nous savions, depuis le DCCT,
qu’optimiser le contrôle glycémique
diminuait la survenue et la progres-
sion des complications microvascu-
laires. EDIC nous montre maintenant
que cette optimisation thérapeutique
“laisse des traces”, avec un bénéfice
– notamment rénal – qui persiste
plusieurs années après la fin de l’in-
tervention optimale. Ces résultats
sont un nouvel encouragement pour
contrôler au mieux, le plus tôt et le
plus longtemps possible nos patients
diabétiques.
M. Joubert,
service d’endocrinologie
et des maladies métaboliques,
CHU Côte-de-Nacre, Caen.
La metformine, traitement
efficace de l’hirsutisme
du SOMPK
Le syndrome des ovaires micro-
polykystiques (SOMPK) est fré-
quemment associé au syndrome
métabolique, dont l’insulinorésis-
tance semble être un déterminant
essentiel. Le lien physiopathologique
exact entre SOMPK et insulinorésis-
tance n’est pas clair à ce jour, mais de
nombreuses équipes ont montré que
la dysfonction ovulatoire du SOMPK
peut être améliorée par les médica-
ments diminuant l’insulinorésistance
(biguanides et thiazolidinediones). En
revanche, l’action de ces substances
sur l’hirsutisme des patientes atteintes
de SOMPK n’avait jamais été préci-
sément évaluée. L’étude prospective
randomisée de Harborne et al. a donc
comparé l’action de la metformine
(1,5 g/j) versus Diane®(1/j, 21 j/mois)
sur l’hirsutisme de 52 patientes
atteintes d’un SOMPK. Diane®est
une pilule contraceptive estropro-
1.
Batterham RL, Cowley MA, Small CJ et
al. Gut hormone PYY3-36 physiologically
inhibits food intake. Nature 2002 ; 418 :
650-4.
2.
Batterham RL, Cohen MA, Ellis SM et al.
Inhibition of food intake in obese subjects by
peptide PYY3-36. N Engl J Med 2003 ; 349 :
941-8.
1.
Steffes M. JAMA 2003 ; 290 : 2159-67.
2.
Steffes M. JAMA 2002 ; 287 : 2563-9.

291
Métabolismes Hormones Diabètes et Nutrition (VII), n° 6, novembre/décembre 2003
Brèves…
Brèves…
gestative associant 35 mg d’éthinyl-
estradiol et 2 mg d’acétate de cypro-
térone, progestatif anti-androgénique
et anti-gonadotrope qui est la molécule
de référence du traitement de l’hirsu-
tisme non tumoral. Les 26 patientes
de chaque bras (metformine/Diane®)
étaient en tout point comparables,
avec notamment un IMC moyen de
32 et un score d’hirsutisme important
(20-22 en moyenne selon Ferriman
et Gallway). Le score d’hirsutisme
ainsi que des paramètres biologiques
d’insulinorésistance et d’hyperandro-
génie ont été réévalués après un an
de l’un ou l’autre des traitements.
De façon non surprenante, la met-
formine a entraîné une diminution
significative de l’index d’insulino-
résistance et de l’IMC, mais n’a pas
eu d’action sur le taux plasmatique
des androgènes ni sur la SHBG (Sex
Hormone Binding Globulin). Diane®
a significativement abaissé les taux
d’androgènes et augmenté le taux de
SHBG, sans aucune action sur l’insu-
linorésistance ou le poids. Les résul-
tats sur l’hirsutisme sont en revanche
troublants : le score de Ferriman et
Gallway a diminué dans les deux
bras, mais davantage sous metformine
que sous Diane®. Le mode d’action
de la metformine sur la régression
de l’hirsutisme ne semble donc pas
médié par la baisse des androgènes
circulants ni par un effet sur l’IGF-I,
dont les taux n’ont pas varié dans
cette étude.
La metformine apparaît comme un
médicament efficace sur l’hirsutisme
du SOMPK, et son mode d’action
est différent de celui des substances
visant à diminuer l’hyperandrogénie.
On attend à présent les études com-
parant la metformine aux fortes
doses d’acétate de cyprotérone, qui
sont utilisées pour le traitement de
l’hirsutisme en combinaison avec
l’estradiol 17b, ainsi que les études
précisant l’effet de l’association des
deux molécules.
M. Joubert
L’hémoglobine glyquée
constitue-t-elle un marqueur
du syndrome métabolique ?
Le syndrome métabolique et son
corollaire, l’insulinorésistance, sont
responsables d’une augmentation
du risque de développer un diabète
et du risque cardiovasculaire, mais on
ne dispose pas d’un marqueur bio-
logique simple pour les identifier.
Osei et al. ont avancé l’hypothèse
que l’hémoglobine glyquée (HbA1C)
pourrait être un tel marqueur dans
une population d’apparentés au pre-
mier degré de diabétiques de type 2
afro-américains de l’Ohio aux États-
Unis. Les auteurs ont étudié les para-
mètres anthropométriques et méta-
boliques suivants, dans un groupe
de 219 sujets non diabétiques : âge,
IMC, rapport taille-hanche, TA sys-
tolique et diastolique, indices d’in-
sulinosecrétion et d’insulinorésis-
tance (minimal model de Bergman
et HOMA), lipides plasmatiques.
Les sujets ont été séparés en trois
groupes correspondant aux tertiles
d’HbA1C suivants : 3,3-4,8 % pour
le 1er tertile, 4,9-5,6 % pour le 2eter-
tile et 5,7-6,4 % pour le 3etertile.
Tous les sujets avaient une glycémie
à jeun inférieure à 110 mg/dl et une
HbA1C normale. La comparaison
des caractéristiques des trois groupes
de patients définis par leur niveau
d’HbA1C a montré que les sujets du
3etertile ont :
– un IMC, un rapport taille-hanche,
une TA systolique et diastolique signi-
ficativement plus élevés que ceux
du 1er et 2etertile ;
– une glycémie à jeun significative-
ment plus élevée (95,5 ± 3,2 mg/dl)
que celle du 1er tertile (83 ± 2,7 mg/dl)
et du 2etertile (78,8 ± 1,5 mg/dl) ;
– une insulinémie et un peptide C
plus élevés (tendance non significa-
tive cependant) ;
– de plus, les sujets du 3etertile ont
un indice Sg d’utilisation du glu-
cose et Si de sensibilité à l’insuline
– calculés par la méthode du minimal
model après test de tolérance au glu-
cose i.v. couplé à une perfusion d’in-
suline exogène – significativement
abaissés par rapport aux sujets des
2eet 3etertile d’HbA1C. De manière
concordante, l’indice HOMA d’insu-
linorésistance des sujets du 3etertile
était plus élevé (3,62 ± 0,26) que
celui des sujets du 1er tertile (2,6 ±
0,21) et du 2etertile d’HbA1C (2,55 ±
0,31). En revanche, l’indice HOMA
évaluant la fonction bsecrétoire
n’était pas différent entre les trois
tertiles d’HbA1C. Ces données ori-
ginales montrent clairement que les
apparentés de diabétiques de type 2
normoglycémiques ayant le niveau
le plus élevé d’HbA1C ont certaines
caractéristiques du syndrome méta-
bolique, et en particulier une insulino-
résistance plus sévère que les sujets
des 1er et 2etertile d’HbA1C. Notons
cependant que les niveaux lipidiques,
en particulier les triglycérides et le
HDL-cholestérol, n’étaient pas diffé-
rents pour les trois tertiles d’HbA1C,
ce qui s’explique, pour les auteurs,
par le phénotype lipidique favorable
des Afro-Américains présentant un
syndrome métabolique, par compa-
raison aux Caucasiens. Ainsi, l’HbA1C
apparaît, dans cette population parti-
culière d’Afro-Américains apparentés
au premier degré de diabétiques de
type 2, comme un marqueur du syn-
drome métabolique. L’extrapolation
de ces données aux sujets d’autres
ethnies, notamment caucasienne, à
des individus ne présentant pas une
prédisposition génétique au diabète
de type 2 nécessitera des études spé-
cifiques, mais on peut déjà retenir
ce marqueur fiable et non coûteux
comme un bon candidat pour repérer
les sujets à risque de développer un
diabète, et donc susceptibles de béné-
ficier des programmes de prévention
hygiéno-diététiques ou d’intervention
pharmacologique par la metformine
comme dans le DPP américain ou le
DPS finlandais.
Y. Reznik,
service d’endocrinologie
et des maladies métaboliques,
CHU Côte-de-Nacre, Caen.
1.
Harborne L. J Clin Endocrinol Metab
2003 ; 88 : 4116-23.
✓
Osei K et al. J Clin Endocrinol Metab 2003 ;
88 : 4596-601.

Brèves…
Brèves…
292
Métabolismes Hormones Diabètes et Nutrition (VII), n° 6, novembre/décembre 2003
PUB
LIPANTYL
Grandeur et misère de la T3
L’étude princeps de Bunevicius et
al. (1), publiée dans le New England
Journal of Medicine en 1999, avait
ouvert le débat sur les effets béné-
fiques de l’utilisation combinée de
T4 et T3 dans le traitement de l’insuf-
fisance thyroïdienne, en particulier
sur la qualité de vie et les fonctions
cognitives. Cette étude aux conclu-
sions originales concernait cepen-
dant un petit échantillon de patients,
comportait un sous-groupe traité par
une dose suppressive d’hormones
thyroïdiennes pour cancer différencié
de la thyroïde, et portait sur une durée
de seulement 5 semaines. Ces biais
méthodologiques entachaient les
conclusions des auteurs, et ont sus-
cité des études méthodologiquement
non criticables. Deux d’entre elles
ont été récemment publiées et vien-
nent infirmer les conclusions opti-
mistes de Bunevicius et al. Walsh et
al. (2) ont mené une étude en double
aveugle, randomisée et contrôlée, sur
une population de 110 patients por-
teurs d’une hypothyroïdie primaire
auto-immune ou idiopathique. Tous
étaient initialement traités par la T4
(dose moyenne, 136 mg/j) et rece-
vaient en cross over au cours de deux
séquences de 10 semaines séparées
par 4 semaines de wash out, soit la
T4 seule, soit T4 + T3 (10 mg de T3
remplacant 50 mg de T4). Les para-
mètres étudiés comportaient une
échelle des symptômes d’insuffi-
sance thyroïdienne, un questionnaire
général de qualité de vie, trois ques-
tionnaires évaluant les fonctions
cognitives et un questionnaire quan-
tifiant les symptômes somatiques,
anxieux, dépressifs et leur retentis-
sement sur la vie sociale. Les patients
sous T4 seule présentaient, à la fin de
l’étude, une TSH significativement
plus basse que ceux sous T4 + T3.
Une seconde analyse a comparé deux
groupes (T4 seule versus T4 + T3)
ayant le même niveau de TSH. La
conclusion des auteurs est l’absence
de tout effet bénéfique du traitement
combiné T4 + T3 sur la qualité de vie,
les fonctions cognitives, l’humeur,
les symptômes d’hypothyroïdie et
la satisfaction subjective du trai-
tement. Sawka et al. (3) ont choisi
d’étudier les effets de la substitution

293
Métabolismes Hormones Diabètes et Nutrition (VII), n° 6, novembre/décembre 2003
Brèves…
Brèves…
par la T3 chez 40 patients hypo-
thyroïdiens ayant des symptomes
dépressifs sous T4 seule. Les patients
recevaient initialement 120 à 132 mg
de T4, et la moitié d’entre eux ont reçu
une demi-dose de T4, compensée par
la prise de 25 mg de T3, alors que
l’autre moitié poursuivait le traite-
ment par T4 seule. L’étude a duré
15 semaines pendant lesquelles les
doses de T4 et T3 ont été ajustées de
manière à maintenir la TSH dans les
valeurs normales. Au terme de l’étude,
les auteurs ont réévalué les scores
d’humeur et constaté l’absence de
toute amélioration des échelles de
dépression sous le traitement com-
biné T4 + T3 par comparaison avec
le groupe maintenu sous T4 seule.
Ces deux études viennent sérieuse-
ment entamer le concept des effets
spécifiques de la T3 sur les fonc-
tions cognitives et l’humeur, et illus-
trent les difficultés méthodologiques
inhérentes à ce type d’étude clinique.
Quant aux effets à plus long terme
de la T3, ils restent à étudier…
Y. Reznik
PUB
LIPANTYL
1.
Bunevicius R. N Engl J Med 1999 ; 340 :
424-9.
2.
Walsh JP. J Clin Endocrinol Metab 2003 ;
88 : 4543-50.
3.
Sawka AM. J Clin Endocrinol Metab
2003 ; 88 : 4551-5.
1
/
5
100%