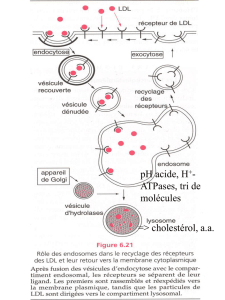
– UE7: Sciences biologiques – Biologie cellulaire

2016-2017 Réception et Transduction du signal Pr. Paumelle
– UE7: Sciences biologiques –
Biologie cellulaire
Indiquer ici, dans cette police s'il y a une annexe en fin d'heure
Semaine : n°1 (du 30/01/17 au
03/02/17)
Date : 30/01/2017
Heure : de 11h00 à
12h00 Professeur : Pr. PAUMELLE
Binôme : n°1 Correcteur : 31
Remarques du professeur Pré-requis pour Hématologie, Immunologie, Enseignements coordonnées.
Le premier ED pour préparer les TP.
2 autres séances d'ED sont basés sur les cours, sur l'analyse de figure, d'articles.
PLAN DU COURS
I. Introduction
II.Comment les cellules communiquent entre elles ?
III. Les principes de la communication cellulaire
1- Transmission du signal
2- Réception du signal
A. Récepteurs nucléaires
B. Récepteurs membranaires
C. Récepteurs canaux
D. Récepteurs couplés aux protéines G
E. Récepteurs enzymes
1/20

2016-2017 Réception et Transduction du signal Pr. Paumelle
I. INTRODUCTION
Les cours magistraux vont nous donner des pistes, des outils que l'on devra utiliser au travers d'expérience,
d'interprétation de résultats pendant les ED/TP.
L'année dernière, on a vu la cellule eucaryote, dans sa globalité, qui est complexe avec une
compartimentation.
Cette année on va prendre du recul pour replacer cette cellule eucaryote au sein d'un organisme, au sein d'un
tissu et essayer de comprendre comment les cellules eucaryotes se comportent, comment elles adaptent leur
comportement cellulaire face à l’environnement.
Objectifs : Toute cellule, au sein d'un organisme multicellulaire, est soumise a des stimuli d'origines diverses
et parfois antagonistes.
→ But : Comprendre les mécanismes moléculaires permettant de définir la réponse cellulaire appropriée
suivant l'environnement (physiologique ou pathologique)
On doit comprendre comment ça se passe.
Les cours de l'an dernier sont importants, ils sont acquis et on va rentrer dans le détail cette année.
Les réponses biologiques principales de la cellule
:
Une cellule a besoin de signaux comme des facteurs de croissance ou des signaux mitogènes pour se
diviser et proliférer.
Elle a également besoin de signaux de survie pour proliférer, sinon on a une induction de la mort cellulaire
(même s'il y a des signaux qui induisent sa prolifération et sa croissance).
Il y aura beaucoup de signaux autour de la cellule pour dicter le comportement cellulaire. La cellule va être
capable de :
–s'orienter vers un programme de prolifération et décroissance cellulaire : quand l'organisme aura
besoin de remplacer des cellules qui meurent.
–Si la cellule n'a pas de signaux de survie : la cellule s'oriente vers l'apoptose.
–Une cellule qui s'est suffisamment divisé peut aussi se différencier. La cellule pourra alors exercer
des choses particulières comme l'inflammation, le métabolisme..
–une cellule peut aussi se déplacer, migrer pour former des structures, des organes.. Elle pourra
aller coloniser un autre tissu (cela dépend des faceturs qui sont présents autour de la cellule).
2/20

2016-2017 Réception et Transduction du signal Pr. Paumelle
→ On a des acteurs moléculaires qui induisent ces changements, ces comportements.
La cellule s'adapte grâce
:
1) Aux récepteurs des signaux environnementaux
La cellule doit recevoir les signaux. Elle ne répond pas à tous les signaux et a absolument besoin de
récepteurs.
Il y a 2 types de récepteurs :
•Récepteurs membranaires
Ils fixent des signaux hydrophiles, qui ne peuvent pas passer la membrane plasmique.
Transmission de l'information dans la cellule puis active une cascade de transduction des gènes.
Ils sont présents au niveau de la membrane plasmique.
•Récepteurs nucléaires
Ils reçoivent aussi des messages.
Ils sont présents soit dans le cytoplasme, soit dans le noyau = dans la cellule.
Ils fixent des signaux hydrophobes qui traversent la membrane plasmique
3/20

2016-2017 Réception et Transduction du signal Pr. Paumelle
Cela induit un engagement dans différents programmes (prolifération, différentiation, migration).
La réponse d'une cellule dépend de la réception du signal via les récepteurs et du contrôle de l'expression
des gènes. L'expression des gènes sera différente en fonction de la réponse biologique qui sera induite.
2) Dépend du contrôle de l'expression des gènes et des protéines :
Cela se fait à différents niveaux de contrôle :
A) Réception et transduction du signal
Au niveau de la cellule, on a des récepteurs qui reçoivent des signaux/Stimulis extra ou intra cellulaire,
conduisant à l'activation ou inactivation d'un gène.
Signaux :
→ Hormones, facteurs de croissance, cytokines
→ Nutriments, toxiques, médicaments
B) Régulation de la transcription des gènes:
Transcription de l'ADN en ARN.
Il peut subir d'autres régulations : génétique, épigénétique, ou rôle des facteurs de transcription.
C) Régulation post-transcriptionnelles sur l'ARNm
→ Régulation de la Maturation, transport ou localisation.
→ Régulation sur sa traduction et sa dégradation.
D) Régulations post-traductionnelles sur la protéine
→ Modifications post-traductionnelles => Dégradation ou activité.
Ce n'est pas parce qu'une protéine est synthétisée qu'elle va jouer un rôle, elle a besoin parfois d'être modifiée
pour être activée.
Ex : phosphorylation pour permettre l'activation.
→ Pour qu'une réponse soit induite, il y a tout un tas de régulations qui agissent.
4/20

2016-2017 Réception et Transduction du signal Pr. Paumelle
Objectif : A la suite des cours, on doit être en mesure :
–savoir nommer les différentes familles de récepteurs et leur mécanisme de transduction du signal →
cours n°1.
–Nommer les différentes protéines impliquées dans les oies de contrôle du cycle cellulaire et de
l'apoptose.
–Définir la fonction des différentes protéines impliquées dans les voies de contrôle du cycle cellulaire
et de l'apoptose.
–Connaître le principe des techniques permettant d'étudier le cycle cellulaire et l'apoptose et leur
application.
–Décrire une figure d'article et l'interpréter.
II.Comment les cellules communiquent entre elles ?
Capable de communiquer au sein de l'organisme.
Il y a plusieurs cas de figure :
Quand les cellules sont
proches les unes des autres :
–les cellules sont proches les unes des autres et pourront faire des pores au niveau de la membrane
plasmique entre elles : ce sont des jonctions communicantes appelées GAP
Ces communications se font en transmettant un second messager. C'est une molécule soluble, très petite.
Elle va passer d'une cellule à une autre, et de cette manière ça va influencer le comportement de la cellule
voisine.
Exemples de 2
nd
messagers : AMP cyclique, des ions, calcium..
Ce sont de toutes petites molécules qui vont être capable de se fixer des enzymes dans la cellule et d'induire
un message à la cellule voisine, pour que la cellule voisine ait le même comportement que la cellule qui est à
côté.
2ème type de communication :
Les cellules sont
proches mais ne sont pas collées : via les molécules d'adhérence responsable de la
reconnaissance et de l'adhérence entre les cellules.
Quand les cellules sont toujours proches mais elles ne sont pas collées : le message peut passer par l'envoie
de signaux à courte distance.
C'est le cas quand les cellules adhèrent les unes aux autres grâce à des protéines d'adhérence, comme les
cathérines (adhérence MEC- cellule) ou les intégrines (cellule-cellule).
Ces protéines d'adhérences permettent de transmettre un signal dans la cellule.
5/20
 6
7
8
9
10
11
12
13
14
15
16
17
18
6
7
8
9
10
11
12
13
14
15
16
17
18
1
/
18
100%