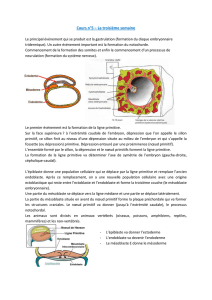
Le rôle du mésoderme somitique dans la

J. Embryol. exp. Morph. Vol. 23,
1,
pp. 109-151, 1970 109
Printed
in Great
Britain
Le rôle du mésoderme
somitique dans la morphogenèse précoce des
membres de l'embryon de Poulet
Par MADELEINE PINOT1
Institut
d''Embryologie
Expérimentale du
C.N.R.S.,
Nogent-sur-Marne,
France
(Directeur :
Professeur Et. Wolff)
La morphogenèse des membres a fait l'objet de nombreux travaux chez
l'embryon de Poulet. Les recherches récentes concernent surtout les processus
intervenant après l'apparition de l'ébauche du membre, et conduisant à sa
morphologie définitive. Le premier stade de développement du membre consiste
en une condensation localisée du mésoderme de la somatopleure. Par la suite
seulement, l'ectoderme sus-jacent acquiert un aspect morphologique particulier:
une crête épaissie à l'extrémité distale. Les causes de la condensation initiale du
mésoderme ne sont pas connues. Le rôle déterminant du constituant mésoder-
mique comme initiateur du développement du membre a été clairement mis en
évidence par Kieny (1959, 1960). Le constituant mésodermique isolé, greffé sous
l'ectoderme du flanc, peut induire la formation d'une crête apicale dans l'ecto-
derme sus-jacent, et provoquer ainsi le développement d'un membre surnumé-
raire.
A quel moment et sous quelles influences le mésoderme acquiert-il cette
propriété
?
Nous nous sommes demandé d'abord à quel stade le territoire du membre
isolé pouvait effectuer son Organogenese de manière 'autonome. Ce problème a
été évoqué par plusieurs auteurs. Hamburger (1938) a greffé dans
le
flanc d'embry-
ons de
3
jours, des bourgeons d'aile et de patte, isolés au stade de 25 à 34 paires
de somites (50-64 h d'incubation). Le tiers environ des greffons
s'est
développé;
les membres obtenus étaient normaux. Des résultats comparables ont été obtenus
récemment par Murillo-Ferrol (1965). L'auteur a greffé dans le flanc, entre aile
et patte, au stade 11 à 18 de Hamburger & Hamilton (1951), des ébauches de
membres de même âge. Le greffon ne donne naissance à un membre normal
que s'il est prélevé sur un embryon du stade 17 (H. & H.) au moins (29 à 32
paires de somites).
Mais on a observé le développement d'un membre à partir de matériel
beaucoup plus
jeune.
(Rudnick, 1942, 1945) a greffé dans le cœlome d'embryons
1 Adresse de routeur: Institut d'Embryologie Expérimentale du C.N.R.S., 49 bis Avenue
de la Belle-Gabrielle, 94-Nogent-sur-Marne, France.

110 M. PINOT
de 3 jours des portions définies de blastoderme, prélevées entre le stade de la
ligne primitive et 15 paires de somites. Des membres reconnaissables, ailes ou
pattes,
se sont développés à partir de fragments de blastoderme de 0 à 5 paires
de somites. Une étude histologique des greffons a révélé que, en plus du squelette
et des muscles, de nombreuses structures se sont formées: tube nerveux,
ganglions, mesonephros, gonade, intestin. C'est donc une partie importante du
blastoderme qui a été greffée. En 1959, Chaube a obtenu la différenciation de
l'aile à partir de territoires prélevés sur des embryons de quatre paires de
somites au moins, et greffés en avant ou en arrière de l'ébauche de l'aile, sur des
embryons de
3
jours.
Aucune
étude
histologique n'a
été
faite. Au stade de quatre
paires de somites, le territoire de l'aile ne peut être isolé de façon précise.
Les auteurs cités ont abouti à des conclusions assez différentes. Ces diver-
gences sont dues sans doute en partie aux techniques diverses utilisées. Mais
elles peuvent s'expliquer également par les difficultés qu'on rencontre pour
isoler précocement le territoire de membre. Les tissus voisins semblent avoir
une influence importante sur le développement du membre. Nous avons repris
le
problème.
Nous avons cherché à savoir
: (i)
à quel stade
le
territoire
de
membre
isolé peut se différencier de manière autonome; (ii) si un tissu voisin, le mesen-
chyme axial, associé à l'ébauche présomptive du membre joue un rôle dans cette
différenciation.
MATERIEL ET METHODES
Nous nous sommes adressé à l'embryon de Poulet
{Gallus gallus)
de la race
Leghorn blanche. Dans une série expérimentale, nous avons utilisé également
l'embryon de Caille
{Coturnix coturnix
Japonica).
Les expériences sont faites sur
des embryons de
7
à
44
paires de somites (stades
9
à
21
de Hamburger
&
Hamil-
ton: 30 h à
3^
jours d'incubation.)
Coupes transversales de blastodermes, au niveau de l'aile,
après prélèvement de l'une des ébauches.
Fig. 1. Embryon de 29 paires de somites. Coupe transversale au niveau de la région
postérieure de l'ébauche de l'aile.
Fig. 2. Embryon de 24 paires de somites. Coupe transversale au niveau de la région
médiane de l'aile. La somatopleure seule est prélevée.
Fig. 3. Embryon de 16 paires de somites. Coupe transversale en arrière du 16e
somite. La pièce intermédiaire est visible. Les trois feuillets embryonnaires sont
excisés; un fragment de la somatopleure est resté en place (flèche).
Fig. 4. Embryon de neuf
paires
de somites. Coupe transversale au niveau présomptif
de l'aile. Le feuillet mésodermique est homogène: rien ne permet de distinguer les
régions somitique, intermédiaire, et latérale. Le prélèvement a été fait
à
petite distance
du tube nerveux.
t.n. = tube nerveux, ch. = chorde, m.s. = mesenchyme somitique,
c.
W.
= canal
de
Wolff,
som. = somatopleure, m.i.
—
feuillet mésodermique indifférencié,/?./. =
pièce intermédiaire. Coloration: glychémalun-éosine.

La morphogenèse précoce des membres 111
&^*4^
100//
I I
+s±i

112 M. PINOT
A. Localisation et prélèvement des ébauches des membres
L'ébauche de l'aile apparaît vers la 50-55ème h de l'incubation, au niveau
des somites
15
à 20, sous forme d'une condensation du mésoderme de la somato-
pleure. Le bourgeon d'aile est parfaitement individualisé au stade de 35 paires de
somites: il se trouve alors au niveau des somites 16 à 20 (Saunders, 1948a).
L'ébauche de la patte est peu visible avant le stade de 30 paires de somites.
Lorsque l'embryon a 35 paires de somites, le bourgeon, au niveau des somites
26 à 32 est aussi développé que celui de l'aile. Le territoire présomptif des mem-
bres a pu être localisé dans une région définie du blastoderme dès le premier
stade de la segmentation somitique.
Le territoire ou
Vébauche
de Vaile a été prélevé aux stades de 7 à 44 paires de
somites. Une étude histologique de 35 embryons de 7 à 30 paires de somites,
fixés après le prélèvement nous a permis d'évaluer les potentialités du territoire
excisé aux différents stades.
Antéropostérieurement: les limites antérieure et postérieure du territoire
présomptif de l'aile ne peuvent être précisées chez des embryons de moins de
15
paires de somites; avant ce stade, elles sont appréciées par référence au dernier
somite formé et au nœud de Hensen, d'après les cartes d'ébauches dues à Wolff
(1936),
Rudnick (1945) et Chaube (1959).
En profondeur: chez les embryons de plus de 23 paires de somites, seule la
somatopleure est prélevée, au niveau de l'aile (fig. 1, 2). Avant ce stade, les
trois feuillets embryonnaires sont
excisés
(fig. 3,4)
;
cette région correspond, pour
le feuillet endodermique, au territoire présomptif de l'intestin grêle (Le Douarin,
1965).
Latéralement: chez les embryons de plus de 15 paires de somites, la limite
entre le mesenchyme somitique, intermédiaire et latéral est nette; au stade de
15 à 20 paires de somites, on excise dans quelques cas, une partie de la pièce
intermédiaire; le plus souvent, au contraire, une frange interne du mésoderme
latéral est laissée en place (fig. 3).
Sur les embryons de moins de 15 paires de somites, le feuillet mésodermique,
au niveau présomptif de l'aile, n'est pas différencié en régions somitique, inter-
médiaire et latérale (fig. 4). On prélève alors un fragment de ce feuillet méso-
dermique indifférencié, avec l'ectoderme et l'endoderme, au ras du tube nerveux
ou à petite distance de celui-ci, au niveau présomptif de l'aile.
Le territoire de la patte est excisé aux stades de 10 à 44 paires de somites.
Situé au niveau des somites 26 à 32, ses limites antéro-postérieures ne peuvent
être précisées avant le stade de 25 paires de somites. La limite entre le méso-
derme latéral et le mésoderme intermédiaire et somitique est nette dans cette
région à partir du stade del
8
à 20 paires de somites. Chez les embryons de
10
à 17
paires de somites, on prélève l'aire présomptif de la patte au ras du tube nerveux.
Notons que d'autres ébauches squelettiques sont présentes dans le méso-
derme de la somatopleure. Différents auteurs (Fell, 1939; Saunders,
1948
A;

La morphogenèse précoce des membres 113
Seno,
1961) s'accordent à localiser l'ébauche sternale, chez les Oiseaux, au
niveau des somites 17^—18 à 21^—22. Le territoire sternal sera donc toujours en
partie prélevé avec le territoire de l'aile. Les côtes se forment au niveau des
somites 19 à 26. Mais nous avons pu montrer (Pinot,
1969 a)
que le matériel
costal, d'origine somitique, ne pénètre dans la plaque latérale qu'après la 50e h
de l'incubation. Le matériel des côtes ne sera donc pas dans le mésoderme
latéral chez les embryons de moins de 22 paires de somites.
B.
Association expérimentale de deux ébauches embryonnaires
(1) Association de deux
territoires
présomptifs de membre
Deux ébauches de l'aile sont associées sur un milieu de culture, de manière
que les deux faces ectodermiques se trouvent à l'extérieur. Elles sont maintenues
accolées pendant 2 à
5
min. Greffé, l'ensemble évolue sans se dissocier. Le milieu
de culture utilisé est le milieu gélose de Wolff & Haffen (1952).
(2) Association de mesenchyme axial et plaque latérale
Pour que les conditions expérimentales soient réalisées, l'association doit
être faite de façon que les rapports entre les deux tissus soient comparables à
ce qu'ils sont dans l'embryon. La surface d'association est donc extrêmement
réduite: elle correspond à l'épaisseur de la somatopleure. Un accolement de
quelques minutes sur un milieu de culture ne suffit pas. Si l'on prolonge la
culture pendant les 8 à 10 h nécessaires pour que l'association soit réalisée, le
développement en greffe est compromis.
Deux techniques ont été utilisées
:
Technique
1 :
association sur un fragment de peau. Un fragment de peau
d'embryon de Poulet est placé sur un milieu de culture, derme en haut; les deux
ébauches sont placées côte à côte sur le derme, auquel elles adhèrent. Les
surfaces à associer sont ainsi maintenues en contact. Après 15 à 20 min sur le
milieu de culture, l'ensemble est greffé.
Technique 2: association en culture de blastoderme. Le blastoderme est cultivé
selon la méthode de Wolff & Simon (1955). Une bande de mesenchyme axial
(intermédiaire et somitique) est prélevée sur un autre embryon et greffée entre
le mesenchyme axial et la plaque latérale ou à la place du mesenchyme axial du
blastoderme, à un niveau choisi. Après 8 à 10 h de culture, le greffon et la
région de la plaque latérale qui lui est associée sont prélevés ensemble et
greffés. C. Greffes cœlomiques
Cette technique a été utilisée pour étudier le développement d'ébauches qui
venaient d'être prélevées, ou d'ébauches associées selon les méthodes précédem-
ment décrites. Une incision est pratiquée à l'aide d'une aiguille de verre dans
la somatopleure en arrière des artères omphalomésentériques, sur un embryon-
hôte de 3 jours d'incubation. Le greffon est introduit dans le cœlome où il est
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
1
/
43
100%