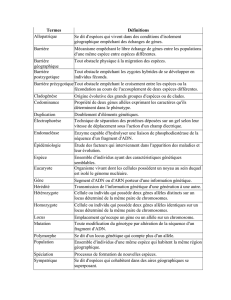
DISCUSSION 1. Analyse de perte d`hétérozygotie 2. Etude de

DISCUSSION
Le présent travail avait pour objectif de tester et de préciser les observations
concernant les sept QTLs associés au cancer mammaire induit chimiquement chez le rat qui
ont été identifiés au laboratoire suite à une analyse de liaison génétique du croisement
impliquant la souche sensible SPRD-Cu3 et la souche résistante WKY/E56. Chacun des sept
locus identifiés a été associé à l'un ou l'autre des trois phénotypes tumoraux à savoir : la
latence tumorale, la multiplicité tumorale et l'agressivité tumorale (Quan et al., 2006). Dans ce
but plusieurs approches ont été suivies.
1. Analyse de perte d’hétérozygotie
La première approche a consisté à s'interroger sur la nature des gènes de
prédisposition : nous nous sommes demandés si ces gènes étaient des gènes suppresseurs de
tumeurs. Nous avons donc recherché des pertes d'hétérozygotie dans les tumeurs, indice
essentiel de la présence de gènes suppresseurs de tumeurs, au niveau de chacun des locus
identifiés statistiquement. Lors de cette analyse aucune perte d’hétérozygotie n’a pu être mise
en évidence dans les sept QTLs identifiés statistiquement. Nous n’avons donc pas pu mettre
en évidence des indices de présence de gènes suppresseurs de tumeurs dans les QTLs. Il est à
noter que de la même manière, aucune perte d’hétérozygotie n’avait été mise en évidence lors
de l’analyse des QTLs Mcs1,Mcs2 et Mcs3 (Shepel et al., 1998). Ceci suggère que la délétion
somatique affectant les gènes suppresseurs de tumeurs potentiellement contenus dans les
régions identifiées n’est pas un évènement qui contribue majoritairement au développement
de cancer mammaire chez le rat. Il ne faut toutefois écarter ni la possibilité que des gènes
suppresseurs de tumeurs soient néanmoins présents dans l’un de ces QTLs ni la possibilité
qu’ils soient impliqués dans la détermination de la sensibilité au cancer mammaire induit au
DMBA. L’inactivation de gènes suppresseurs de tumeurs peut être due à d’autres évènements
tels que des modifications épigénétiques.
2. Etude de lignées congéniques
La deuxième approche a consisté dans un premier temps à mesurer l’implication
individuelle de six des sept QTLs identifiés dans le contrôle de la sensibilité au cancer
mammaire induit au carcinogène DMBA et dans un deuxième temps, à évaluer les effets
d’une combinaison génétique et tester les relations d'épistasie entre les QTLs localisés sur les
50

chromosomes 5 et 18.
Ces études nous ont permis d’assurer l’existence de cinq des sept QTLs de sensibilité
au cancer mammaire chimiquement induit identifiés au laboratoire : les locus associés au
phénotype de multiplicité tumorale localisés sur les chromosomes 1p.tel et 5, le locus associé
au phénotype de latence tumorale localisé sur le chromosome 9 et les locus associés au
phénotype de vitesse de croissance tumorale localisés sur les chromosomes 10 et 18 (en
collaboration avec D. Stieber pour les QTL chromosomes 5, 9, 18). Nous avons également
précisé l’implication de ces locus dans le contrôle de chacun des trois phénotypes tumoraux.
2.1 Phénotype de multiplicité tumorale
Nous avons pu montrer que les QTLs localisés sur les chromosomes 1p.tel, 5, 10 et 18
identifiés statistiquement manifestent leur effet sur le phénotype de multiplicité tumorale au
moins en partie indépendamment de la présence des autres QTLs identifiés par les analyses de
liaisons. L’allèle d’origine WKY du locus 1p.tel, locus que nous avons appelé Mcstm3, est un
allèle cryptique et est capable à lui seul d’augmenter la sensibilité au cancer mammaire induit
chimiquement. Cette augmentation était attendue sur base des études de liaisons génétiques. A
l’inverse, les allèles d’origine WKY de chacun des trois autres locus localisés sur les
chromosomes 5, 10 et 18 sont capables, à eux seuls de réduire de façon significative le risque
de développer des tumeurs mammaires induites au carcinogène DMBA.
Le locus identifié sur le chromosome 5 a été appelé Mcstm1 et la présence de l’allèle
WKY à ce locus induit à elle seule une réduction de la multiplicité tumorale de 65% en
comparaison à la lignée parentale SPRD-Cu3. Le QTL Mcstm1 est situé dans la région bordée
par le marqueur D5Rat124 (19.2 Mb) et le gène Pla2g2a (157.6 Mb) (Figure 17.A). Etant
donné la taille importante de cette région (138.4 Mb), il est probable qu’elle contienne
plusieurs QTLs distincts. Cette région est homologue à une région du chromosome 4 de souris
qui contient deux QTLs (Mmtg1 et Mmtg2) associés à la masse tumorale mammaire induite
par un transgène (middle T gene polyome) (Le Voyer et al., 2000). D’autre part, l’intervalle
que nous avons défini comprend les QTLs Mcs5a1, Mcs5a2,Mcs5b et Mcs5c (sous-QTLs de
Mcs5) associés au phénotype de multiplicité tumorale lors de l’analyse du croisement WF X
WKY. L’étude de ces QTLs a permis de mettre en évidence que les QTLs Mcs5a1 et Mcs5b
sont cryptiques et qu’il existe des interactions d’épistasie entre les deux locus Mcs5a et
Mcs5b (Lan et al., 2001; Samuelson et al., 2005; Samuelson et al., 2007). A l’instar du QTL
Mcs5,Mcstm1 pourrait donc contenir plusieurs QTLs pouvant agir dans des sens opposés. Et
compte tenu du fait qu’il a été montré que la structure haplotypique de la région Mcs5b est
commune aux lignées SPRD-Cu3 et WF et que celle-ci est différente de celle de la lignée
51

WKY, un des sous-QTLs de Mcstm1 pourrait contenir des gènes contrôlant potentiellement la
sensibilité au cancer mammaire qui seraient communs à Mcs5 et se trouveraient dans Msc5b
(Stieber et al., 2007). La région du QTL Mcstm1 se superpose également au QTL Emca1
associé à la sensibilité au cancer mammaire induit par oestrogènes. L’analyse de la structure
haplotypique de la région commune dans les différentes lignées impliquées dans les
croisements a permis de conclure que dans le cas ou ces QTLs sont identiques, les gènes
causaux se trouveraient dans une région distale de celle du QTL Mcs5 (en fait deux régions
ont été mises en évidence) (Stieber et al., 2007). Ce dernier argument appuie l’hypothèse de
l’existence de sous-QTLs dans la région Mcstm1.
Les lignées congéniques C10.Mcsta1 et C18.Mcstm2/Mcsta2 montrent respectivement
une réduction de 18% et 33% de la multiplicité tumorale. De manière remarquable, les QTLs
localisés sur les chromosomes 10 et 18 avaient été associés dans un premier temps au
phénotype de vitesse de croissance tumorale. L’observation d’un phénotype de multiplicité
tumorale de ces lignées congéniques intégrant une région initialement associée au phénotype
de vitesse de croissance peut s’expliquer dans le cas de la lignée congénique C.10.Mcsta1 par
le fait que la réduction, bien que significative de la multiplicité tumorale est modeste et de ce
fait difficile à détecter lors de l’analyse de liaison génétique. Dans le cas de la lignée
congénique C18.Mcstm2/Mcsta2, une explication de ce fait est que la région qui contient le
QTL d’agressivité présente également un QTL de multiplicité mais à un niveau
statistiquement faible qui avait été ignoré en première analyse (Quan et al., 2006; Stieber et
al., 2007). Les résultats de l’analyse phénotypique de la lignée congénique
C.18.Mcstm2/Mcsta2 démontrent que la liaison génétique qui n’était que faiblement
suggestive est bien réelle et nous avons appelé ce QTL Mcstm2. Le QTL Mcstm2 est le
premier locus de sensibilité au cancer mammaire induit chimiquement assigné au
chromosome 18. Il faut toutefois noter qu’un locus associé au cancer mammaire induit par
œstrogènes, Emca2, a été localisé sur le chromosome 18 (Gould et al., 2004) (Figure 17.B).
Plusieurs arguments permettent de soutenir l’hypothèse que les deux QTLs sont distincts,
notamment l’analyse de la structure haplotypique de la région commune aux deux locus qui a
révélé que les lignées étudiées ne partagent pas d’haplotype commun au niveau de la région
d’intérêt (Stieber et al., 2007).
En ce qui concerne le locus 1q.tel, la lignée congénique associée ne montre aucune
réduction de la multiplicité tumorale en comparaison avec la lignée parentale sensible. Nous
n’avons donc pas été en mesure de confirmer les résultats des analyses de liaisons qui
prédisaient que la région du chromosome 1 bordée par les marqueurs D1Rat324 (229.5 Mb) et
D1Rat88 (265.8 Mb) contenait un locus impliqué dans la détermination de la multiplicité
tumorale. Ces observations n’excluent pas la possibilité que le QTL 1q.tel joue un rôle dans la
52

détermination de la sensibilité au cancer mammaire. Une hypothèse pour expliquer la perte du
phénotype lors de la réalisation de la lignée congénique est que des interactions intergéniques
peuvent être nécessaires pour permettre à un QTL donné d’exercer son effet sur le phénotype
(phénomène d’épistasie, comme observé sur le QTL Mcsa2 sur le chromosome 18).
2.2 Phénotype de latence tumorale
Nous avons montré que les QTLs de sensibilité au cancer mammaire identifiés
statistiquement sur les chromosomes 5, 9 et 10 manifestent leur effet sur le phénotype de
latence tumorale au moins en partie indépendamment de la présence des autres QTLs
identifiés. En ce qui concerne le QTL localisé sur le chromosome 9 que nous avons appelé
Mcstl1, les analyses de liaison prédisaient l’effet inverse de celui observé lors de la
caractérisation phénotypique. Dans ce dernier cas c’est l’allèle WKY qui augmente le temps
de latence et confère la résistance.
De manière surprenante, bien qu’aucun QTL associé à ce trait n’avait été détecté dans
les régions définissant les lignées congéniques C5.Mcstm1 et C10.Mcsta1, celles-ci montrent
des différences significatives en terme de latence tumorale par rapport à la lignée parentale
sensible. Les allèles d’origine WKY de chacun des deux locus sont capables, à eux seuls,
d’augmenter et de réduire, respectivement, de façon significative le temps d’apparition de la
première tumeur. Le QTL Mcstm1 (chromosome 5) est donc impliqué à la fois dans la
détermination de la multiplicité et de la latence tumorale, l’allèle WKY de Mcstm1 conférant
la résistance dans les deux cas. Le QTL Mcsta1 localisé sur le chromosome 10 est également
impliqué dans la détermination des deux traits mais dans ce cas, l’allèle WKY confère soit la
résistance soit la sensibilité. Ces résultats constituent un argument pour appuyer l’hypothèse
que les trois phénotypes tumoraux sont au moins en partie indépendants.
2.3 Phénotype de vitesse de croissance tumorale
Nous avons pu montrer que le QTL localisé sur le chromosome 10 identifié
statistiquement manifeste son effet sur le phénotype de vitesse de croissance tumorale au
moins en partie indépendamment des autres QTLs identifiés. La congénique C10.Mcsta1
montre une réduction statistiquement significative de 26% de la vitesse de croissance des
tumeurs les plus agressives par rapport à celle de la lignée parentale. L’allèle d’origine WKY
du locus est capable à lui seul de diminuer la vitesse de croissance de tumeurs mammaires
induites chimiquement. Cette diminution était attendue sur base des études de liaisons
génétiques. Nous avons appelé ce QTL, Mcsta1. Le locus Mcsta1 est donc impliqué dans
53

chacun des trois phénotypes tumoraux. Cependant, son effet est cryptique lorsque l’on
considère le phénotype de latence tumorale. A l’inverse, en terme de multiplicité et de vitesse
de croissance, l’allèle WKY confère la résistance mais l’amplitude de l’effet du QTL sur le
premier est réduite (18% de réduction de la charge tumorale) par rapport à celle de l’effet sur
la vitesse de croissance.
2.4 Evaluation des effets d’une combinaison génétique entre les QTLs localisés sur les
chromosomes 5 et 18.
Lors de l’analyse des relations d’épistasie entre les locus localisés sur les
chromosomes 5 (Mcstm1) et 18 (initialement appelé Mcstm2) nous avons pu observer que la
lignée double congénique présentait une réduction de 49% du taux moyen de croissance de la
tumeur la plus rapide par rapport à celui de la lignée parentale. Rappelons qu’aucune des deux
lignées congéniques simples ne présente de réduction de la vitesse de croissance tumorale par
rapport à la lignée sensible. Les allèles WKY des QTLs localisés sur les chromosomes 5 et 18
interagissent de manière synergique pour moduler la vitesse de croissance tumorale. Ces
résultats constituent également une confirmation physique du fait que le QTL détecté sur le
chromosome 18 par les analyses de liaisons est impliqué dans la vitesse de croissance
tumorale, bien que son effet ne puisse être détecté dans la congénique simple du chromosome
18 (Quan et al., 2006; Stieber et al., 2007). Des interactions épistatiques semblables avaient
été mises en évidence par M.Gould et collaborateurs lors de l’analyse du croisement entre les
lignées WKY et WF. Ces analyses ont montré que bien que le locus Mcsm1 (« Mammary
cancer susceptibility modifier1 », RNO6) n’exerce pas à lui seul d’effet majeur sur le
phénotype de multiplicité tumorale, l’allèle WKY au locus diminue l’effet de l’allèle de
résistance WKY au locus Mcs8 (RNO14) (notons que dans ce cas-ci, l’allèle d’origine WKY
du locus Mcs8 est capable à lui seul de diminuer la multiplicité tumorale) (Lan et al., 2001).
La région du chromosome 18 contenant le QTL Mcstm2 contrôle à la fois la
multiplicité tumorale et la vitesse de croissance tumorale. Cependant les deux traits sont
indépendamment modulés par la région du chromosome 5 contenant le QTL Mcstm1, étant
donné que l’allèle WKY du QTL Mcstm2 n’a pas d’effet sur la multiplicité en présence de
l’allèle WKY du QTL Mcstm1 alors que ces deux régions agissent de manière synergique sur
la vitesse de croissance tumorale. Les phénotypes de multiplicité tumorale et de croissance
tumorale sont au moins en partie indépendants. Ceci justifie que nous ayons considéré que la
région du chromosome 18 bordée par les marqueurs D18Wox8 (32.5 Mb) et D18Rat44 (86.9
Mb) contient deux QTLs et décidé d’utiliser une dénomination distincte pour chacun d’eux
(Mcstm2 et Mcsta2).
De manière remarquable, aucun QTL n’avait encore été associé au phénotype de
54
 6
7
8
9
10
6
7
8
9
10
1
/
10
100%