Stabilité et variabilité des génomes et évolution

(6 semaines)
S
Stabilité et variabilité des génomes
tabilité et variabilité des génomes
et évolution
et évolution
Rappels:
Questions des pages 84 et 85
Les mutations modifient la séquence de la molécule ADN, et sont responsables de l’apparition de
nouveaux allèles. Il y a trois types de mutations: substitution, addition et délétion.
De plus il existe un lien entre le phénotype d’un individu et son génotype.
1
1 L'apport de l'étude des génomes : les innovations génétiques.
L'apport de l'étude des génomes : les innovations génétiques.
Quelles sont les conséquences des mutations sur les gènes ?
1.1
1.1 Le polymorphisme des gènes.
Le polymorphisme des gènes.
TP1 : Le polymorphisme des gènes.
1.1.1
1.1.1 Les conséquences des mutations par substitution.
Les conséquences des mutations par substitution.
Substitution : remplacement d'un nucléotide par un autre.
Les mutations silencieuses n'entraînent aucune modification dans la séquence des acides aminés du
polypeptide codé par l'allèle concerné par suite de
la redondance du code génétique.
Les mutations non-sens transforment un
triplet de nucléotides codant pour un acide aminé
en un codon STOP. Cela entraîne la production
d'un polypeptide tronqué, généralement non
fonctionnel.
Les mutations faux-sens, qui se traduisent par
le remplacement d'un acide aminé par un autre
dans la séquence polypeptidique, ont des
conséquences variables : certaines (conservatrices)
ne modifient pas les propriétés du polypeptide,
alors que d'autres (non conservatrices) les
affectent.
1.1.2
1.1.2 Les conséquences des mutations par addition ou délétion.
Les conséquences des mutations par addition ou délétion.
Addition (ou insertion) : insertion d'un nouveau nucléotide.
Délétion : perte d'un nucléotide.
Les insertions et les délétions entraînent un décalage du cadre de lecture et, généralement, la mise en
place d'une séquence d'acides aminés modifiée à partir de la délétion ou de l'addition ; le plus souvent, le
polypeptide codé par cet allèle est non fonctionnel.
1.1.3
1.1.3 Conclusion.
Conclusion.
La mutation d'un gène peut conduire à l'apparition d'un nouvel allèle pour ce gène.
Un gène est qualifié de polymorphe si au moins deux de ses allèles sont présents dans l’espèce à une
fréquence supérieure ou égale à 1 %.
Au sein d’une espèce, le polymorphisme des séquences d'ADN résulte de l’accumulation de
mutations (substitution, addition ou délétion d’un ou de plusieurs nucléotides) ont des
conséquences phénotypiques variables.
1

Remarque : seule les mutations germinales (cellules souches des gamètes) sont susceptibles de se
transmettre à la descendance. Les mutations somatiques, quant à elles ne se transmettent pas.
1.2
1.2 Les familles de gènes.
Les familles de gènes.
Comment expliquer la ressemblance entre certains gènes ?
TP2 : Les familles de gènes + « Evolmol »
1.2.1
1.2.1 Les mécanismes à l'origine des familles de gènes.
Les mécanismes à l'origine des familles de gènes.
La duplication génique conduit à la formation de deux nouveaux gènes identiques. Ces deux gènes
subissent alors des mutations et divergent.
Ainsi la vision des couleurs qui repose sur trois gènes codant pour trois pigments ayant des spectres
d'absorption différents repose sur une double duplication d'un gène ancestral.
Ces gènes ont ensuite subi des translocations car deux gènes sont localisés sur le chromosome X et
le troisième est localisé sur le chromosome 7.
1.2.2
1.2.2 Les familles de gènes et les innovations évolutives.
Les familles de gènes et les innovations évolutives.
On considère que plus il y a de similitudes entre deux gènes appartenant à la même famille de gènes,
plus récente est la duplication.
Il est parfois possible de préciser le moment de la duplication. En effet, tous les singes de l'Amérique
du Sud ne voient que deux couleurs alors que les autres primates en voient trois, ce qui situe la dernière
duplication après la séparation des deux lignées.
La duplication génique est donc un mécanisme essentiel de la complexification des génomes.
1.2.3
1.2.3 Conclusion.
Conclusion.
Au sein du génome d’une espèce, les similitudes entre gènes (familles de gènes) sont
interprétées comme le résultat d’une ou plusieurs duplications d’un gène ancestral.
La divergence des gènes d’une même famille s’expliquent par l’accumulation de mutations.
Dans certains cas, ces processus peuvent conduire à l’acquisition de gènes correspondant à de
nouvelles fonctions.
Les innovations génétiques sont aléatoires et leur nature ne dépend pas des caractéristiques
du milieu.
Exercices 2 page 101 et 5 page 102
2
divergence
gène
ancestral
duplication
gène
ancestral
mutations
conservatrices
fonction
différente
nouveau
gène deux gènes
homologues
fonction
conservée
mutations non
conservatrices
même
famille de
gènes

2
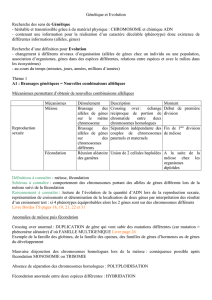
2 Méiose et fécondation participent à la stabilité de l'espèce.
Méiose et fécondation participent à la stabilité de l'espèce.
TP 3 : Les cycles de développement.
Le Maintien du caryotype qui est indispensable à la
stabilité de l'espèce se fait grâce à la méiose et à la
fécondation :
–la méiose produit des cellules haploïdes (n
chromosomes) à partir de cellules diploïdes (2n
chromosomes),
–la fécondation rétablit la phase diploïde par la
fusion de cellules sexuelles haploïdes.
Chez les organismes présentant une
reproduction sexuée, une phase haploïde et une
phase diploïde alternent.
2.1
2.1 La méiose permet la formation de cellules haploïdes.
La méiose permet la formation de cellules haploïdes.
TP4 : La méiose.
2.1.1
2.1.1 La première division de méiose.
La première division de méiose.
Elle est précédée d'une duplication des chromosomes. En prophase I, il y a appariement des
chromosomes homologues.
Les chromosomes homologues migrent vers les pôles opposés de la cellule, ainsi, la disjonction des
chromosomes à l'anaphase I permet le passage à la phase haploïde.
Les deux cellules filles obtenues possèdent n chromosomes à deux chromatides.
2.1.2
2.1.2 La deuxième division de méiose.
La deuxième division de méiose.
Les chromatides de chaque chromosomes se séparent à l'anaphase II comme dans une mitose
normale, mais elle concerne une cellule haploïde.
Les deux cellules filles obtenues possèdent n chromosomes à une chromatide.
2.1.3
2.1.3 Conclusion.
Conclusion.
La méiose assure le passage de la phase diploïde à la phase haploïde. Elle suit une phase de
réplication de l'ADN et se compose de deux divisions cellulaires successives qui conduisent à la
présence d’un lot haploïde de chromosomes par cellule fille.
Visualisation de la vidéo « meiose.mov ».
2.2
2.2 La fécondation rétablit la diploïdie.
La fécondation rétablit la diploïdie.
2.2.1
2.2.1 Les conséquences de la fécondation.
Les conséquences de la fécondation.
Pages 114 et 115.
La fécondation est l'union d'un gamète mâle à n chromosomes et d'un gamète femelle à n
chromosomes. La cellule œuf obtenue possède donc 2n chromosomes.
2.2.2
2.2.2 Des anomalies lors de la méiose aboutissent à des anomalies
Des anomalies lors de la méiose aboutissent à des anomalies
chromosomiques.
chromosomiques.
Questions 1 à 3 page 117.
Si lors de la première division de méiose, les chromosomes d'une même paire migrent vers le même
pôle cellulaire, alors les cellules filles auront un caryotype anormal.
Ainsi, après la fécondation, la cellule œuf qui héritera du matériel génétique d'un gamète anormal
3

sera porteuse d'une anomalie chromosomique :
–une trisomie dans le cas où une paire est constitué trois chromosomes au lieu de deux,
–une monosomie dans le cas où une paire est constitué d'un seul chromosome au lieu de deux.
Remarque : les conséquences sont variables selon la paire concernée, en effet, cela varie de la non
viabilité à des malformations plus ou moins graves.
2.2.3
2.2.3 Conclusion.
Conclusion.
La fécondation rétablit la diploïdie en réunissant les lots haploïdes des gamètes d’une même
espèce. Mais des perturbations dans la répartition des chromosomes lors de la formation des
gamètes conduisent à des anomalies du nombre des chromosomes.
Exercices 3 page 127 et 5 page 128
3
3 Méiose et fécondation sont à l'origine du brassage génétique.
Méiose et fécondation sont à l'origine du brassage génétique.
3.1
3.1 De nombreux gènes chez l'individu sont hétérozygotes.
De nombreux gènes chez l'individu sont hétérozygotes.
Question 1 page 131
Dans le cas des cellules diploïdes, chaque gène est représenté par deux allèles. On dit de l'individu
qu'il est hétérozygote s'il possède deux allèles différents pour un gène donné et qu'il est homozygote s'il
possède deux allèles identiques pour un gène donné.
Cette hétérozygotie résulte de l'existence au sein d'une espèce de nombreux allèles pour un même
gène. Un individu diploïde est hétérozygote pour 6 à 7% de ses gènes.
La variabilité allélique se manifeste au sein de l’espèce par une hétérozygotie à de nombreux
loci.
3.2
3.2 Le brassage génétique chez une espèce diploïde.
Le brassage génétique chez une espèce diploïde.
TP 5 : Le brassage génétique chez une espèce diploïde.
Rappels : Les allèles d'un gène peuvent être dominants, récessifs ou codominants.
–
un allèle dominant s'exprime même s'il est seul,
–
deux allèles récessifs doivent être présents pour s'exprimer,
–
deux allèles codominants s'expriment en même temps.
Conventions d'écriture : soit deux gènes a et b dont les allèles a+ et b+ sont dominants (sauvage).
Le phénotype sauvage s'écrira [a+,b+].
Le génotypes correspondant s'écrira (a+//a+,b+//b+) dans le cas d'un homozygote.
3.2.1
3.2.1 La transmission d'un gène.
La transmission d'un gène.
Questions 1 et 2 page 133.
Le croisement entre deux individus homozygotes pour le gène étudié, un « sauvage » et un « muté »
aboutit à une génération F1 uniquement constituée d'individus de phénotype « sauvage » hétérozygotes.
Les individus issus d'un test-cross (croisement entre individus F1 et individus double récessif) seront
alors constitués de 50% de phénotype « sauvage » et de 50% de phénotype « muté ».
Un croisement-test permet ainsi de déterminer les gamètes produits par l'individu donc son génotype.
3.2.2
3.2.2 La Transmission de deux gènes.
La Transmission de deux gènes.
Si les individus issus d'un test-cross n'ont pas les proportions 50% - 50%, cela signifie que deux gènes
interviennent. Il existe alors deux possibilités :
–si les quatre types de gamètes sont en quantité égale (résultat du test-cross), cela signifie que les
deux gènes sont non liés, c'est à dire sur deux chromosomes non homologues, dans ce cas on parle de
brassage interchromosomique,
–si les quatre types de gamètes sont en quantité différentes, cela signifie que les deux gènes sont
liés, c'est à dire sur le même chromosome, dans ce cas on parle de brassage intrachromosomique.
4

3.2.3
3.2.3 Conclusion.
Conclusion.
La variabilité génétique est due aux brassages interchromosomiques lors de la méiose.
Cette variabilité est accrue par le brassage intrachromosomique lors de la méiose.
Cette variabilité génétique est également liée à la réunion au hasard des gamètes lors de la
fécondation.
3.3
3.3 Le brassage génétique chez une espèce haploïde.
Le brassage génétique chez une espèce haploïde.
TP 6 : Le brassage chromosomique chez un champignon : Sordaria macrospora.
Chez un organisme haploïde, quand un caractère correspond à un seul gène, alors le phénotype
correspond nécessairement à un allèle unique.
Dans le cas de Sordaria, on croise une souche sauvage (spores noire) avec une souche mutée (spores
jaunes). Après la fécondation (caryogamie), il se forme des périthèces dans lesquelles se forment des
asques. Les asques contiennent des spores issue de la méiose suivie d'une mitose, ce qui explique
qu'elles soient deux par deux.
Les asques produites sont :
–de type I (4-4) brassage interchromosomique.
–de type II (2-2-2-2) ou (2-4-2) brassage
intrachromosomique.
Le brassage interchromosomique est dû à la migration
indépendante des chromosomes homologues de chaque paire
lors de l'anaphase de la première division de méiose.
Le brassage intrachromosomique, ou recombinaison
homologue par crossing-over, a lieu entre chromosomes
homologues appariés lors de la prophase de la première
division de méiose.
Exercice sordaria
Une faible fréquence de crossing-over indique que le gène est proche du centromère.
Exercice 5 page 146
4
4 Étude de trois exemples de relations entre mécanismes de
Étude de trois exemples de relations entre mécanismes de
l’évolution et génétique.
l’évolution et génétique.
Les innovations génétiques peuvent être favorables, défavorables ou neutres pour la survie
de l’espèce.
4.1
4.1 Mutations et avantage sélectif.
Mutations et avantage sélectif.
Questions 1 et 4 page 149
Une innovation génétique, comme un nouvel allèle, peut conférer à l'individu qui la reçoit, une
probabilité de survie plus grande que celle des organismes qui ne la possèdent pas.
Les mutations qui confèrent un avantage sélectif aux individus qui en sont porteurs ont une
probabilité plus grande de se répandre dans la population au fur et à mesure des générations. Ceci
est un processus de sélection naturelle positive.
En revanche, l'innovation génétique qui aura pour effet de diminuer la capacité reproductive des
organismes est soumise à une sélection naturelle négative, et tend à être éliminée.
Remarque : Parmi les innovations génétiques seules celles qui affectent les cellules
germinales d’un individu peuvent avoir un impact évolutif.
Questions 2 à 4 page 151
La fréquence de certains allèles est forte dans les populations alors que les individus homozygotes qui
5
brassage
interchromosomique
= phénotype parental
brassage
intrachromosomique
= phénotype recombiné
 6
6
1
/
6
100%