Histones désacétylases et signalisation estrogénique dans le cancer du sein

Correspondances en Métabolismes Hormones Diabètes et Nutrition - Vol. XV - n° 8 - octobre 2011
284
dossier thématique
Épigénétique : nouvelle donne
en endocrinologie
Histones désacétylases et signalisation
estrogénique dans le cancer du sein
Histone deacetylases and estrogen signalling in breast cancer
Nathalie Boulle*,**, Aurélien Linares*, Marion Lapierre*, Vincent Cavailles*
Points forts
Highlights
»
Les récepteurs des estrogènes (RE) régulent la transcription
de gènes spécifiques en interagissant avec des complexes
multiprotéiques qui contiennent des histones acétyltransférases
(HAT) et des histones désacétylases (HDAC).
»
Les RE et leurs corégulateurs transcriptionnels peuvent être
modifiés par acétylation sur les résidus lysine. Cette modification
post-traductionnelle contrôle la capacité des RE à interagir avec
l’ADN et à moduler la transcription.
»
Différentes HDAC sont recrutées directement ou indirectement
par les RE et régulent l’activité de ces récepteurs ainsi que leur
expression dans les cellules cancéreuses mammaires.
»
Les HDAC sont impliquées à plusieurs niveaux dans la signalisation
estrogénique ; leur expression est dérégulée dans les tumeurs
du sein.
»
Les inhibiteurs de HDAC ont conduit à de premiers résultats
prometteurs dans le traitement des cancers du sein avancés,
en association avec une chimiothérapie ou avec un traitement
hormonal.
Mots-clés : Récepteur des estrogènes – Histone désacétylase – Cancer
du sein – Inhibiteur d’histone désacétylase – Thérapie hormonale.
The estrogen receptor (ER) regulates transcription of specific
genes by interacting with multiprotein complexes containing
histone acetyltransferase (HAT) and histone deacetylase
(HDAC).
RE and their transcriptional coregulators can be modified
by acetylation on lysine residues. This post-translational
modification controls their ability to interact with DNA and
modulate transcription.
Different HDAC are recruited directly or indirectly by the RE
and regulate the activity and the expression of these receptors
in breast cancer cells.
HDAC are involved at several levels in estrogen signaling and
their expression is deregulated in breast tumors.
HDAC inhibitors led to promising results in the treatment of
advanced breast cancer in combination with chemotherapy
or hormonal treatments.
Keywords: Estrogen receptors – Histone deacetylase – Breast
cancer – Histone deacetylase inhibitor – Hormone therapy.
* Institut de recherche
en cancérologie de
Montpellier ; Inserm, U896,
Montpellier ; université
Montpellier-1 ; CRLC Val-
d’Aurelle-Paul-Lamarque,
Montpellier.
** Laboratoire de biologie
cellulaire et hormonale,
CHRU Arnaud-de-
Villeneuve, Montpellier.
Signalisation par les estrogènes
et cancer du sein
Le cancer du sein est un problème majeur de santé
publique (lire le rapport de l’INCa sur la situation du
cancer en France en 2010 : http://www.e-cancer.fr). Les
estrogènes jouent un rôle clé dans la physiologie et la
cancérogenèse mammaires. Ils agissent par liaison à
des récepteurs spécifiques appartenant à la famille des
récepteurs nucléaires, les récepteurs des estrogènes (RE)
α et β. Environ 70 à 80 % des cancers du sein infiltrants
expriment le REα, ce qui a permis la mise en place de
traitements visant à bloquer la signalisation estrogé-
nique (1). Cependant, ces thérapies sont limitées par
des phénomènes de résistance apparaissant d’emblée,
ou au cours du traitement (2).
Au niveau moléculaire, la liaison de l’hormone
aux RE induit une modification conformationnelle,
entraînant leur fixation sur des promoteurs de gènes
estrogénosensibles et le recrutement de complexes
multiprotéiques (3). Ces complexes sont constitués
de différents corégulateurs transcriptionnels (activa-
teurs ou répresseurs) possédant pour la plupart des
activités enzymatiques. Par exemple, de nombreuses
histones acétyltransférases (HAT) et histones désacé-
tylases (HDAC) sont recrutées par les RE et agissent
sur la structure de la chromatine afin de moduler la
transcription des gènes cibles.

Correspondances en Métabolismes Hormones Diabètes et Nutrition - Vol. XV - n° 8 - octobre 2011
285
Histones désacétylases et signalisation estrogénique dans le cancer du sein
HDAC et inhibiteurs de HDAC
Chez l’homme, 18 HDAC ont été identifiées, classées
en 4 familles (I à IV) selon leur degré d’homologie de
séquences et la nature de leur domaine catalytique
(utilisant le zinc ou le NAD comme cofacteur) [4]. Les
HDAC sont impliquées dans divers mécanismes cellu-
laires généraux tels que la prolifération cellulaire ou
l’apoptose mais également dans la différenciation de
tissus spécifiques, notamment pour les HDAC de classe II
(5). Par ailleurs, de nombreux travaux ont montré qu’au-
delà des histones, les HDAC avaient des substrats variés
(facteurs de transcription, enzymes, cytosquelette, etc.).
Les premiers inhibiteurs de HDAC ciblant le domaine
catalytique zinc-dépendant ont été développés dès les
années 1970 (6). Très rapidement, leur intérêt en tant
qu’agents antitumoraux a été mis en évidence dans les
tumeurs hématologiques et solides, ce qui a conduit
à développer de nouveaux inhibiteurs de structure et
d’efficacité variées (7). À ce jour, une centaine d’essais
cliniques utilisant ces molécules, seules ou en asso-
ciation avec d’autres traitements, sont en cours dans
le domaine du cancer (8).
HDAC et cancer du sein
Dans le cancer du sein, une diminution significative du
niveau d’acétylation des histones entre le tissu mam-
maire normal et les carcinomes mammaires in situ a
été mise en évidence, ce qui suggère que les anomalies
d’acétylation des protéines constituaient un événement
précoce de la cancérogenèse mammaire. Des études
réalisées au niveau ARNm et/ou protéines dans des
tumeurs du sein invasives ont mis en évidence la pré-
sence de certaines HDAC (principalement HDAC 1, 2, 3 et
6) dans les cellules tumorales mammaires, associée pour
certaines d’entre elles à une valeur pronostique dans
les tumeurs du sein (9). De plus, l’analyse de données
issues de puces à ADN a confirmé une dérégulation
d’expression de plusieurs HDAC dans les cancers du
sein par rapport au tissu mammaire normal (10). Enfin,
l’HDAC 6 a été impliquée dans la mobilité des cellules
tumorales mammaires ainsi que dans leur prolifération
indépendante du support.
Les données récentes montrent que les HDAC et leurs
inhibiteurs régulent de manière très complexe la signa-
lisation par les estrogènes dans les cellules de cancer
du sein. Deux revues récentes (10, 11) développent
les différents niveaux de contrôle, détaillés plus loin
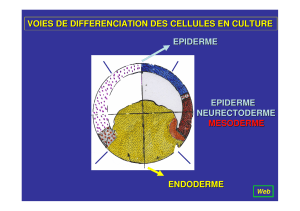
et illustrés dans la figure.
HDAC et contrôle de l’activité
et de l’expression des RE
Par la technique d’immunoprécipitation de chroma-
tine (ChIP), différentes HDAC de classe I ou II ont été
Figure. HDAC et signalisation estrogénique.
Activité
transcriptionnelle
AAAAA
HDAC
Stabilité de la protéine
Stabilité ARNm ?
Transcription
REα
REα
REα
miR ?
Gène REα
ADN
MEF2
Ac
Ac Ac
Ac
CoF
REαAc
HDAC
DNMT
Hsp90 Hsp90

Correspondances en Métabolismes Hormones Diabètes et Nutrition - Vol. XV - n° 8 - octobre 2011
286
dossier thématique
Épigénétique : nouvelle donne
en endocrinologie
mises en évidence sur des promoteurs cibles des RE,
en présence de ligands agonistes ou antagonistes.
L’association directe de certaines HDAC (HDAC 1,
4, 5, 6 et 9) avec plusieurs domaines du REα a été
démontrée par des expériences d’interaction in vitro
et par co-immunoprécipitation. Les HDAC peuvent
aussi être recrutées de façon indirecte, par interaction
avec les corégulateurs transcriptionnels, qui lient le
récepteur en présence d’agonistes ou d’antagonistes
estrogéniques. Plusieurs HDAC appartenant à des
classes différentes ont été impliquées dans la régula-
tion de l’activité transcriptionnelle des RE (10). Ainsi,
la surexpression de HDAC 1 inhibe l’activité trans-
criptionnelle estradiol (E2)-dépendante du REα. Une
approche par siRNA dans les cellules cancéreuses du
sein T47D a également montré le rôle de HDAC 2 dans
la régulation de l’expression et de l’activité du REα.
Par ailleurs, une étude récente du laboratoire d’Olson
révèle que les HDAC de classe II agissent comme des
modulateurs importants du REα (12). En effet, HDAC 5
et HDAC 9 répriment l’activité transcriptionnelle du
REα et participent à la régulation de son promoteur.
L’HDAC 7, une autre HDAC de classe II, semble jouer
un rôle unique dans la répression E2-dépendante de
l’expression des gènes. Par ailleurs, l’HDAC 4 régule
négativement la transactivation du REα en présence
d’E2 ou d’anti-estrogènes, et l’HDAC 6 participe aux
effets non génomiques des estrogènes en intera-
gissant avec la forme membranaire du REα. Enfin,
concernant les HDAC de classe III, une étude a montré
que l’invalidation du gène SIRT1 était associée à une
diminution de l’expression du REα.
Acétylation des RE
et corégulateurs transcriptionnels
Le REα et plusieurs corégulateurs transcriptionnels (par
exemple RIP140/NRIP1, TIF2 ou PGC1α) sont des pro-
téines acétylées, ce qui illustre la complexité des effets
pouvant résulter de la modulation du niveau d’acéty-
lation en réponse à l’inhibition des HDAC (10). Cette
modification post-traductionnelle contrôle différents
paramètres tels que la stabilité et la localisation sub-
cellulaire de la protéine, ou encore sa capacité à inter-
agir avec d’autres molécules (ADN, ligands ou autres
protéines). Deux groupes ont décrit l’acétylation du
REα sur différentes lysines (K266/K268 et K302/K303)
situées dans le domaine de liaison à l’ADN et dans la
région charnière de la protéine. L’acétylation des lysines
K302/303 semble diminuer la réponse du REα au ligand,
et celle des résidus K266/268 augmente la liaison à
l’ADN du récepteur et son activité de transactivation
ligand-dépendante. Cependant, les enzymes impliquées
sont peu connues (rôles de p300 et SIRT1 démontrés
in vitro) et le rôle de cette modification reste encore
relativement flou.
Inhibiteurs de HDAC
et signalisation estrogénique
Dans différents modèles cellulaires exprimant le REα
(sein, endomètre, ovaire, etc.), le traitement par inhibi-
teurs de HDAC à large spectre (TSA, SAHA ou LBH589)
conduit à une diminution très marquée de l’expression
du REα au niveau de l’ARNm et de la protéine (13). Le
même type de régulation est obtenu avec le sirtinol
(inhibiteur des HDAC de classe III), mais pas avec des
inhibiteurs sélectifs des HDAC de classe II. Les méca-
nismes impliqués sont variés et font intervenir des
régulations à la fois transcriptionnelles et post-trans-
criptionnelles. De manière intéressante, les HDAC 5 et
9 inhibent la transcription du gène REα par compéti-
tion sur son promoteur avec le facteur de transcrip-
tion MEF2. Différents micro-ARN pourraient également
intervenir, car plusieurs d’entre eux ciblant l’ARNm REα
sont régulés par les inhibiteurs de HDAC. La diminu-
tion d’expression du REα sous l’effet d’inhibiteurs de
HDAC semble également d’origine post-traductionnelle,
impliquant notamment la protéine chaperon Hsp90
(l’hyper acétylation de Hsp90 diminue son association
avec le REα, conduisant ainsi à l’ubiquitination et à la
dégradation du récepteur) ou l’ubiquitine ligase CHIP
(C-terminal Hsc70 interacting protein). Enfin, les HDAC
modulent l’activité transcriptionnelle des RE. En effet,
dans des modèles de cellules cancéreuses humaines
transfectées de manière stable avec un gène rappor-
teur piloté par un élément de réponse aux estrogènes,
l’inhibition des HDAC augmente le potentiel de trans-
activation des REα et des REβ et fait apparaître une
activité agoniste des anti-estrogènes liée à l’inhibition
de l’expression du REα.
Inhibiteurs de HDAC
et réponse aux anti-estrogènes
Un point important en termes de retombées théra-
peutiques potentielles concerne l’effet des inhibiteurs
de HDAC sur la réponse de cellules cancéreuses mam-
maires aux anti-estrogènes (11). Le traitement de cel-
lules MCF-7, sensibles ou insensibles au tamoxifène, par
un inhibiteur de HDAC (FK228) inhibe la prolifération

Correspondances en Métabolismes Hormones Diabètes et Nutrition - Vol. XV - n° 8 - octobre 2011
286
dossier thématique
Épigénétique : nouvelle donne
en endocrinologie
mises en évidence sur des promoteurs cibles des RE,
en présence de ligands agonistes ou antagonistes.
L’association directe de certaines HDAC (HDAC 1,
4, 5, 6 et 9) avec plusieurs domaines du REα a été
démontrée par des expériences d’interaction in vitro
et par co-immunoprécipitation. Les HDAC peuvent
aussi être recrutées de façon indirecte, par interaction
avec les corégulateurs transcriptionnels, qui lient le
récepteur en présence d’agonistes ou d’antagonistes
estrogéniques. Plusieurs HDAC appartenant à des
classes différentes ont été impliquées dans la régula-
tion de l’activité transcriptionnelle des RE (10). Ainsi,
la surexpression de HDAC 1 inhibe l’activité trans-
criptionnelle estradiol (E2)-dépendante du REα. Une
approche par siRNA dans les cellules cancéreuses du
sein T47D a également montré le rôle de HDAC 2 dans
la régulation de l’expression et de l’activité du REα.
Par ailleurs, une étude récente du laboratoire d’Olson
révèle que les HDAC de classe II agissent comme des
modulateurs importants du REα (12). En effet, HDAC 5
et HDAC 9 répriment l’activité transcriptionnelle du
REα et participent à la régulation de son promoteur.
L’HDAC 7, une autre HDAC de classe II, semble jouer
un rôle unique dans la répression E2-dépendante de
l’expression des gènes. Par ailleurs, l’HDAC 4 régule
négativement la transactivation du REα en présence
d’E2 ou d’anti-estrogènes, et l’HDAC 6 participe aux
effets non génomiques des estrogènes en intera-
gissant avec la forme membranaire du REα. Enfin,
concernant les HDAC de classe III, une étude a montré
que l’invalidation du gène SIRT1 était associée à une
diminution de l’expression du REα.
Acétylation des RE
et corégulateurs transcriptionnels
Le REα et plusieurs corégulateurs transcriptionnels (par
exemple RIP140/NRIP1, TIF2 ou PGC1α) sont des pro-
téines acétylées, ce qui illustre la complexité des effets
pouvant résulter de la modulation du niveau d’acéty-
lation en réponse à l’inhibition des HDAC (10). Cette
modification post-traductionnelle contrôle différents
paramètres tels que la stabilité et la localisation sub-
cellulaire de la protéine, ou encore sa capacité à inter-
agir avec d’autres molécules (ADN, ligands ou autres
protéines). Deux groupes ont décrit l’acétylation du
REα sur différentes lysines (K266/K268 et K302/K303)
situées dans le domaine de liaison à l’ADN et dans la
région charnière de la protéine. L’acétylation des lysines
K302/303 semble diminuer la réponse du REα au ligand,
et celle des résidus K266/268 augmente la liaison à
l’ADN du récepteur et son activité de transactivation
ligand-dépendante. Cependant, les enzymes impliquées
sont peu connues (rôles de p300 et SIRT1 démontrés
in vitro) et le rôle de cette modification reste encore
relativement flou.
Inhibiteurs de HDAC
et signalisation estrogénique
Dans différents modèles cellulaires exprimant le REα
(sein, endomètre, ovaire, etc.), le traitement par inhibi-
teurs de HDAC à large spectre (TSA, SAHA ou LBH589)
conduit à une diminution très marquée de l’expression
du REα au niveau de l’ARNm et de la protéine (13). Le
même type de régulation est obtenu avec le sirtinol
(inhibiteur des HDAC de classe III), mais pas avec des
inhibiteurs sélectifs des HDAC de classe II. Les méca-
nismes impliqués sont variés et font intervenir des
régulations à la fois transcriptionnelles et post-trans-
criptionnelles. De manière intéressante, les HDAC 5 et
9 inhibent la transcription du gène REα par compéti-
tion sur son promoteur avec le facteur de transcrip-
tion MEF2. Différents micro-ARN pourraient également
intervenir, car plusieurs d’entre eux ciblant l’ARNm REα
sont régulés par les inhibiteurs de HDAC. La diminu-
tion d’expression du REα sous l’effet d’inhibiteurs de
HDAC semble également d’origine post-traductionnelle,
impliquant notamment la protéine chaperon Hsp90
(l’hyper acétylation de Hsp90 diminue son association
avec le REα, conduisant ainsi à l’ubiquitination et à la
dégradation du récepteur) ou l’ubiquitine ligase CHIP
(C-terminal Hsc70 interacting protein). Enfin, les HDAC
modulent l’activité transcriptionnelle des RE. En effet,
dans des modèles de cellules cancéreuses humaines
transfectées de manière stable avec un gène rappor-
teur piloté par un élément de réponse aux estrogènes,
l’inhibition des HDAC augmente le potentiel de trans-
activation des REα et des REβ et fait apparaître une
activité agoniste des anti-estrogènes liée à l’inhibition
de l’expression du REα.
Inhibiteurs de HDAC
et réponse aux anti-estrogènes
Un point important en termes de retombées théra-
peutiques potentielles concerne l’effet des inhibiteurs
de HDAC sur la réponse de cellules cancéreuses mam-
maires aux anti-estrogènes (11). Le traitement de cel-
lules MCF-7, sensibles ou insensibles au tamoxifène, par
un inhibiteur de HDAC (FK228) inhibe la prolifération

Correspondances en Métabolismes Hormones Diabètes et Nutrition - Vol. XV - n° 8 - octobre 2011
287
Histones désacétylases et signalisation estrogénique dans le cancer du sein
tumorale des cellules et abolit également la résistance
au tamoxifène. Cette resensibilisation ne semble pas
limitée au tamoxifène : elle est également observée avec
les inhibiteurs d’aromatase. Les mécanismes impliqués
dans cette resensibilisation aux thérapies hormonales
sont encore peu connus et pourraient impliquer le blo-
cage de l’activation de PAK1, qui joue un rôle dans la
signalisation par les facteurs de croissance.
Réexpression du REα
par les inhibiteurs de HDAC
Le second point important concerne la régulation de
l’expression du REα dans les cellules cancéreuses mam-
maires qui n’expriment pas ce gène. Différentes études
ont en effet montré que le traitement de ces cellules par
la TSA ou par l’acide valproïque (VPA) permettait la réex-
pression de l’ARNm du Reα, et ce, de manière synergique
avec des inhibiteurs de l’ADN méthyltransférase tels que
la 5-aza-2’-désoxycytidine. La réactivation de l’expres-
sion du gène REα est associée à une réorganisation de
la structure de la chromatine au niveau du promoteur et
semble couplée à la perte du récepteur à l’EGF (EGFR).
L’association de la TSA et du 5-aza-dC, un inhibiteur
des DNMT, restaure la sensibilité au tamoxifène dans la
lignée cellulaire mammaire MDA-MB 235 REα négative.
D’autres mécanismes peuvent être impliqués : diminu-
tion de l’expression d’EGFR et des voies de signalisation
associées, ou encore augmentation de l’expression de
l’aromatase.
Conclusion
L’analyse des liens entre HDAC et RE met en évidence
des niveaux d’interaction multiples et complexes, et
des effets opposés sur l’expression du REα dans les
cellules cancéreuses mammaires exprimant ou non
le récepteur. Plusieurs questions importantes restent
à résoudre afin de mieux apprécier ces dialogues au
niveau transcriptionnel : quels sont les rôles précis des
différentes isoformes des HDAC ? Les HDAC régulent-
elles la signalisation estrogénique indépendamment de
leur activité catalytique ? Quel est leur rôle exact dans
les tumeurs du sein résistantes aux thérapies hormo-
nales ? Malgré ces questions qui restent encore sans
réponse, les inhibiteurs des HDAC ont conduit à de
premiers résultats très prometteurs dans le traitement
des cancers du sein avancés, en association avec une
chimiothérapie ou un traitement hormonal (tamoxifène
ou inhibiteurs de l’aromatase) [14]. Plusieurs essais cli-
niques sont en cours dans ce domaine. Cependant, des
études complémentaires seront nécessaires pour définir
les meilleures associations thérapeutiques ainsi que les
biomarqueurs permettant d’identifier les patientes les
mieux à même de bénéficier de ces nouvelles thérapies
combinatoires (15).
■
1.
McGuire WL, Horwitz KB, Pearson OH, Segaloff A. Current
status of estrogen and progesterone receptors in breast cancer.
Cancer 1977;39:2934-47.
2.
Massarweh S, Schiff R. Unraveling the mechanisms of endo-
crine resistance in breast cancer: new therapeutic opportunities.
Clin Cancer Res 2007;13:1950-4.
3.Heldring N, Pike A, Andersson S et al.
Estrogen receptors:
how do they signal and what are their targets. Physiol Rev
2007;87:905-31.
4.Yang XJ, Seto E. HATs and HDACs: from structure, function
and regulation to novel strategies for therapy and prevention.
Oncogene 2007;26:5310-8.
5.Haberland M, Montgomery RL, Olson EN. The many roles of
histone deacetylases in development and physiology: impli-
cations for disease and therapy. Nat Rev Genet 2009;10:32-42.
6.Minucci S, Pelicci PG. Histone deacetylase inhibitors and
the promise of epigenetic (and more) treatments for cancer.
Nat Rev Cancer 2006;6:38-51.
7.Witt O, Deubzer HE, Milde T, Oehme I. HDAC family: what
are the cancer relevant targets? Cancer Lett 2009;277:8-21.
8.
Tan J, Cang S, Ma Y, Petrillo RL, Liu D. Novel histone dea-
cetylase inhibitors in clinical trials as anti-cancer agents.
J Hematol Oncol 2010;3:5.
9.Weichert W. HDAC expression and clinical prognosis in
human malignancies. Cancer Lett 2009;280:168-76.
10.Linares A, Dalenc F, Balaguer P, Boulle N, Cavailles V.
Manipulating protein acetylation in breast cancer: a pro-
mising approach in combination with hormonal therapies?
J Biomed Biotechnol 2011;2011:856985.
11.Thomas S, Munster PN. Histone deacetylase inhibitor
induced modulation of anti-estrogen therapy. Cancer Lett
2009;280:184-91.
12.Van Rooij E, Fielitz J, Sutherland LB et al.
Myocyte enhancer
factor 2 and class II histone deacetylases control a gender-spe-
cific pathway of cardioprotection mediated by the estrogen
receptor. Circ Res 2010;106:155-65.
13.
Margueron R, Duong V, Castet A, Cavailles V. Histone dea-
cetylase inhibition and estrogen signalling in human breast
cancer cells. Biochem Pharmacol 2004;68:1239-46.
14.Munster PN, Thurn KT, Thomas S et al.
A phase II study of
the histone deacetylase inhibitor vorinostat combined with
tamoxifen for the treatment of patients with hormone the-
rapy-resistant breast cancer. Br J Cancer 2011;104:1828-35.
15.
Stimson L, La Thangue NB. Biomarkers for predicting clini-
cal responses to HDAC inhibitors. Cancer Lett 2009;280:177-83.
Références
 6
6
1
/
6
100%