Les mécanismes du déclenchement pubertaire : signaux périphériques, hypothalamiques et environnementaux

Act. Méd. Int. - Métabolismes - Hormones - Nutrition, Volume V, n° 3, mai-juin 2001
101
La puberté normale
La puberté et, secondairement, la mise en
place de la fonction de reproduction sont deux
événements majeurs qui caractérisent la vie
de nombreuses espèces, y compris celle de
l’homme. Ces événements sont la consé-
quence de l’activation de l’axe hypothalamo-
hypophyso-gonadique avec augmentation
progressive de la production de stéroïdes
gonadiques, responsables des changements
physiologiques observés à cette période de la
vie. L’hypothalamus est l’élément cardinal de
cet axe, où se trouvent localisés les neurones
à GnRH. Ces neurones, après migration pen-
dant la vie embryonnaire de la placode olfac-
tive vers l’hypothalamus, projettent leurs ter-
minaisons axonales vers l’éminence médiane.
Le développement mammaire chez la fille et
l’augmentation du volume testiculaire chez le
garçon sont les premiers témoins cliniques du
développement pubertaire apparaissant à un
âge moyen de 10,5 ans et 11,5 ans respecti-
vement. Il existe cependant une importante
variabilité physiologique de 8 ans à 13,5 ans
chez la fille et de 9 ans à 14 ans chez le gar-
çon. En deçà et au-delà de ces âges, on parle
respectivement de puberté précoce et de
puberté tardive. Récemment, une large étude
menée par des pédiatres praticiens américains
concernant plus de 17 000 filles a rapporté
un âge moyen de 8,9 ans pour l’apparition
des seins chez les filles noires et 10,0 ans chez
les filles blanches (1). Parmi celles-ci, 15 %
avaient un développement mammaire apparu
avant 8 ans (et 48 % chez les Noires).
Cependant, l’âge moyen de la ménarche, sur-
venant à 12,7 ans, n’est pas particulièrement
avancé et relativement similaire à l’âge
moyen rapporté antérieurement par Tanner et
Davies, qui est de 12,6 ans. Ces données sug-
gèrent un début plus précoce du développe-
ment mammaire qui semble être associé à une
Les mécanismes du déclenchement pubertaire :
signaux périphériques, hypothalamiques
et environnementaux
M.C. Lebrethon*, J.P. Bourguignon*
* Service de pédiatrie ambulatoire et médecine de l’adolescent, CHU Sart. Tilman, université de
Liège, Belgique.
✎Il existe une grande variabilité phy-
siologique de l’âge où survient la
puberté.
✎ Dans de nombreuses espèces y
compris chez l’homme, le déclenche-
ment pubertaire est précédé par une
accélération de la fréquence de la
sécrétion pulsatile de Gonadotropin
Releasing Hormone (GnRH) au niveau
hypothalamique.
✎Deux acides aminés sont particuliè-
rement dominants dans le contrôle de
la sécrétion pulsatile de GnRH : le glu-
tamate qui a un rôle stimulateur et
l’acide
γ
-aminobutyrique (GABA) qui
a un rôle inhibiteur.
✎Les cellules astrogliales jouent un
rôle important dans la régulation de
la sécrétion pulsatile de GnRH notam-
ment, par la production de
Transforming Growth Factor
α
(TGF
α
).
✎Les mécanismes génétiques ont
probablement une implication dans
les variations de l’âge du déclenche-
ment pubertaire.
✎Une des facettes possibles du rôle
des stéroïdes sexuels est le dimorphis-
me sexuel dans le déclenchement de
la puberté.
✎La leptine est un facteur permissif
possible du déclenchement pubertaire.
✎Les xénoestrogènes pourraient être
impliqués dans la survenue de puberté
précoce chez les enfants adoptés.
Mise au point

progression plus lente de la puberté. Dans les
pays autres que les États-Unis, aucune étude
montrant une telle avance du développement
mammaire n’a été rapportée. Aux Pays-Bas,
un travail réalisé en 1997 (2) montre des âges
moyens du début de développement mam-
maire et de la ménarche à 10,7 et 13,1 ans, et
comparables aux données établies quelque
20 ans plus tôt. Chez la fille, en dehors du
développement mammaire, le développement
de la pilosité pubienne et axillaire est essen-
tiellement dû à la sécrétion d’androgènes sur-
rénaliens. L’apparition précoce de ce déve-
loppement avant l’âge de 8 ans peut témoi-
gner d’une adrénarche prématurée indépen-
damment de la mise en route de la puberté.
Les mécanismes
du déclenchement pubertaire
Il est classiquement admis que le déclenche-
ment pubertaire est précédé d’une accéléra-
tion de la fréquence de la sécrétion pulsatile
de GnRH, et secondairement, de LH au
niveau hypophysaire. Chez l’homme, cette
accélération commence par apparaître au
cours du sommeil et connaît des variations
circadiennes caractéristiques de la période
pubertaire. Plus tard, c’est surtout l’augmen-
tation de l’amplitude de la sécrétion de GnRH
et de LH qui détermine le cycle ovulatoire et
la fonction de reproduction chez la femme.
Les recherches menées pour élucider les
mécanismes responsables du déclenchement
pubertaire, que ce soit dans les variations
physiologiques ou les formes pathologiques
de pubertés précoce et tardive, n’ont pas
abouti à une réponse univoque. Des signaux
importants ont été isolés aussi bien à partir de
l’environnement extérieur qu’à partir de
structures périphériques, tels les gonades ou
le tissu adipeux, et, enfin, au sein de l’hypo-
thalamus lui-même
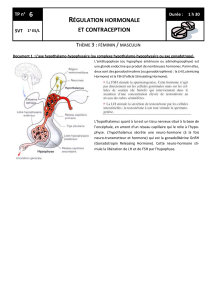
(figure 1)
.
Signaux hypothalamiques
La notion de générateur de pulsatilité fut
introduite par Knobil (3) sur la base de
décharges électrophysiologiques périodiques
enregistrées dans l’hypothalamus. L’exis-
tence d’un générateur de pulsatilité intrin-
sèque aux neurones à GnRH fut proposée
pour la première fois sur la base de la
démonstration d’une sécrétion pulsatile de
GnRH à partir de culture de lignées cellu-
laires de neurones à GnRH immortalisés, les
cellules GT1 (4). Plus récemment, chez le
singe (5) et chez le rat (6),une sécrétion pul-
satile de GnRH a été mise en évidence à par-
tir de culture d’explants de la placode olfactive,
confirmant la présence d’un générateur de
pulsatilité propre aux neurones à GnRH.
Act. Méd. Int. - Métabolismes - Hormones - Nutrition, Volume V, n° 3, mai-juin 2001
102
Figure 1. Signaux périphériques, hypothalamiques et environnementaux intervenant dans la régulation de
l’axe hypophyso-gonadique.
aut
auto-t
o-test
est
Vrai ou faux ?
1. Une augmentation d’amplitude de la sécrétion pulsatile de GnRH précède
le début pubertaire.
2. Les neurones GABAergiques ont un rôle stimulateur de la sécrétion pulsatile
de GnRH.
3. En cas d’insuffisance gonadique primitive, les gonadotrophines sont plus
basses chez la fille que chez le garçon.
4. Le TGFαest capable d’entraîner la maturation sexuelle aussi bien chez l’ani-
mal que chez l’homme.
Réponses page 116
Mise au point
n
n°
°1
1

Act. Méd. Int. - Métabolismes - Hormones - Nutrition, Volume V, n° 3, mai-juin 2001
103
L’existence d’un réseau neuronal proche des
terminaisons des axones à GnRH et capable
d’induire une sécrétion pulsatile au niveau de
ces derniers a largement été évoquée dans la lit-
térature, notamment dans nos travaux chez le
rat. C’est ce que nous appelons le “générateur
de pulsatilité extrinsèque” aux neurones à
GnRH. Bien que la composition de ce “généra-
teur de pulsatilité extrinsèque” ne soit pas
encore parfaitement élucidée, nous envisa-
geons au moins trois composants possibles
dans sa structure, dont deux acides aminés qui
semblent particulièrement dominants dans dif-
férentes espèces. D’une part, le glutamate est
un acide aminé excitateur majeur capable de
stimuler la sécrétion de GnRH, notamment via
les récepteurs au N-méthyl-D-aspartate
(NMDA) aussi bien chez le singe (7) que chez
le rat (8). D’autre part, l’acide γ-amino buty-
rique (GABA) exerce un rôle inhibiteur fonda-
mental (9). Un troisième composant, important
dans l’hypothalamus du rat, est le rétrocontrôle
négatif exercé sur les récepteurs NMDA par la
GnRH1-5,produit de dégradation de la GnRH1-10
sous l’action d’une prolylendopeptidase (PEP)
(10). Chez le rat, l’ontogenèse de la sécrétion
pulsatile de GnRH implique une accélération
significative de la sécrétion pulsatile de GnRH,
avant la puberté, surtout entre le 15eet le
25ejour postnatal. Cette accélération prépuber-
taire implique trois processus : une diminution
de l’action inhibitrice exercée par le GABA via
les récepteurs de type GABAA,une diminution
du rétrocontrôle négatif exercé par la GnRH1-5
au niveau des récepteurs de type NMDA qui
exprime la sous-unité NR2A et une augmenta-
tion de la biosynthèse de glutamate à partir de
la glutamine (11). Chez le singe, Terasawa
considère la levée d’inhibition exercée par les
neurones GABAergiques comme un élément
critique dans les mécanismes du déclenche-
ment pubertaire (12).
Chez l’homme, le rôle des acides aminés,
glutamate et GABA, a été illustré par la des-
cription d’une puberté précoce chez une
fillette d’un an avec hyperglycinémie non
cétosique. La glycine qui stimulait excessive-
ment certains des récepteurs NMDA où elle
exerce une activité co-agoniste du glutamate,
a sans doute entraîné, via ce mécanisme, une
puberté précoce qui a régressé suite à l’admi-
nistration de divers anti-épileptiques ago-
nistes des récepteurs GABA (13).
À côté des neurones sécrétant les acides ami-
nés excitateurs ou inhibiteurs, les cellules
astrogliales, en contact étroit avec les neu-
rones à GnRH notamment au niveau de l’émi-
nence médiane, produisent de nombreux fac-
teurs de croissance capables de stimuler la
sécrétion de GnRH. Parmi ceux-ci, citons
deux membres de la famille des facteurs de
croissance épidermiques, le TGFαet les neu-
régulines. Ces facteurs de croissance stimu-
lent indirectement la sécrétion de GnRH par
liaison à des récepteurs de type tyrosine kina-
se de la famille erbB, localisés sur les astro-
cytes (14). Cette capacité du TGFαà stimuler
la maturation sexuelle chez l’animal a trouvé
son corollaire clinique dans deux observa-
tions de puberté précoce avec hamartomes.
L’analyse histologique de ces tumeurs a mon-
tré une expression importante du TGFα,et
non de la GnRH, comme cela avait été pro-
posé antérieurement pour expliquer cette
pathologie (15). Ce sont aussi les cellules
astrogliales qui sécrètent la glutamine synthé-
tase, une enzyme capitale pour transformer le
glutamate en glutamine qui sera elle-même
régulée en glutamate par l’action de la gluta-
minase, au sein des neurones glutamater-
giques. Nous avons observé que le pouvoir
sécrétagogue de la glutamine augmente très
tôt au cours de l’ontogenèse et contribue ainsi
au contrôle de la pulsatilité.
De plus en plus de travaux récents s’intéres-
sent à la recherche de gènes impliqués dans le
déclenchement de la puberté. Parmi ces tra-
vaux, citons ceux de Wierman et al. qui
démontrent l’implication de facteurs de trans-
cription, Brn-4, SCIP et SOX-2 dans l’ex-
pression du gène de la GnRH (16). Ojeda et
1. Élévation postnatale des gonadotrophines plus marquée
chez la fille que chez le garçon agonadique.
2. Après l’activation postnatale, entrée en “pause juvénile”
à 6 mois chez le garçon et 2-3 ans chez la fille.
3. Entrée en puberté chez la fille (M2) à un âge moyen
inférieur de 0,8 ans par rapport au garçon (G2).
4. Maturations des variations circadiennes de la sécrétion
pulsatile de LH à la puberté, plus rapide chez la fille que
chez le garçon.
5. Incidence considérablement plus grande de la puberté
précoce chez la fille que chez le garçon.
6. Incidence nettement plus grande de la puberté tardive
chez le garçon.
7. Puberté précoce freinée plus aisément chez le garçon
avec des agonistes de GnRH moyennement actifs.
8. Puberté induite plus aisément chez la fille
suite à l’administration pulsatile de GnRH.
9. Pic de croissance pubertaire avancée (de type féminin)
chez les sujets XY avec insensibilité aux androgènes.
Tableau I. Observations cliniques en faveur d’une freination de l’axe hypothalamo-hypophyso-gonadique
moindre chez la fille que chez le garçon au cours du développement.

Act. Méd. Int. - Métabolismes - Hormones - Nutrition, Volume V, n° 3, mai-juin 2001
104
al. ont également montré que deux gènes
homéobox, Oct-2 et TTF-1, pouvaient être
impliqués relativement précocement dans la
cascade des événements qui conduisent au
déclenchement pubertaire (14). Le gène Oct-2
serait un facteur de transcription impliqué
notamment dans l’expression de TGFα. Ces
travaux ouvrent la porte sur la matérialisation
des mécanismes génétiques certainement
impliqués dans les variations de l’âge du
déclenchement pubertaire.
Signaux périphériques
De nombreuses données cliniques et expéri-
mentales établissent un lien entre nutrition,
puberté et reproduction. Dès 1974, Frisch et
al. (17) suggéraient qu’une masse critique doit
être atteinte pour permettre l’apparition des
premières règles. Chez des jeunes femmes
comme chez l’homme, un jeûne est capable
d’induire une inhibition de la sécrétion pulsa-
tile de LH, même à court terme (18). Chez le
rat et le singe, la malnutrition aiguë ou chro-
nique inhibe également la sécrétion gonado-
trope qui est restaurée suite à la reprise de
l’alimentation (19, 20). La leptine identifiée
suite au clonage du gène ob en 1994 (21) est
apparue très rapidement comme un signal
d’information sur le statut énergétique entre la
périphérie et le système nerveux central. Cette
protéine, synthétisée et sécrétée par les cel-
lules adipeuses, exerce principalement ses
effets au niveau hypothalamique en inhibant
les signaux orexigènes, dont le plus puissant
est le neuropeptide Y (NPY) et en stimulant
les signaux anorexigènes, dont le Cocaine-
amphetamine-regulated transcript (CART).
La leptine s’est révélée être également impli-
quée dans la régulation de l’axe hypothalamo-
hypophyso-gonadique. Les souris ob/ob, défi-
cientes en leptine, présentent en plus de leur
obésité un hypogonadisme, lequel est corrigé
par un traitement à la leptine (22). Les rares
observations de déficience en leptine chez
l’homme indiquent un hypogonadisme à l’âge
adulte
(23)
et une puberté avancée dans le cas
d’une jeune patiente traitée avec de la leptine
(24). Nos travaux ont montré que la leptine
accélère la sécrétion pulsatile de GnRH chez
le rat prépubère de 15 jours (25) et que cette
action serait partiellement transmise par le
CART (26). En ce qui concerne le NPY, les
travaux sont plus divergents, suggérant soit
une inhibition, soit une stimulation de la
sécrétion de GnRH. Chez le singe mâle, Plant
et al. (27) considèrent le NPY comme un fac-
teur inhibiteur de la sécrétion pulsatile de
GnRH durant la période prépubertaire. Chez
le singe femelle, Terasawa et al. (28) montrent
cependant une stimulation par le NPY de la
sécrétion de GnRH. Chez le rat, les travaux
d’Aubert et al. (29) plaident plus en faveur
d’un rôle inhibiteur du NPY dans le mécanis-
me de maturation sexuelle alors que nos
propres travaux in vitro montrent une accélé-
ration de la sécrétion pulsatile de GnRH par le
NPY dans l’hypothalamus prépubère et cela
indépendamment des effets de la leptine (25).
Ces résultats a priori divergents peuvent trou-
ver quelques explications dans l’action chro-
nique ou aiguë du NPY et l’environnement en
stéroïdes sexuels présents ou absents (ani-
maux intacts ou gonadectomisés).
Parmi les autres signaux nutritionnels, il est
important de souligner le rôle possible de
l’insuline, du glucose et de l’IGF1 dans ce
processus de maturation sexuelle (30).
Les stéroïdes sexuels ont été parmi les pre-
miers messagers périphériques impliqués
dans le rétrocontrôle inhibiteur de la sécrétion
de GnRH au niveau de ce qui fut baptisé le
“gonadostat” hypothalamique. Une des
facettes possibles de l’effet des stéroïdes
sexuels est le dimorphisme sexuel dans le
déclenchement de la puberté. Un faisceau
d’arguments cliniques indique une prompti-
tude à l’entrée en puberté qui est relativement
plus grande chez la fille et une inertie plus
grande chez le garçon
(tableau I, p. 103)
.
L’étude du dimorphisme sexuel de l’axe
hypothalamo-hypophyso-gonadique au cours
du développement chez le singe donne lieu à
des observations en tout point comparables à
celles obtenues chez l’homme. La gonadecto-
mie néonatale entraîne des concentrations
élevées de FSH (et de LH) qui se réduisent
avec l’âge, mais plus tard et dans une moindre
mesure chez la femelle, et réaugmentent à un
âge plus précoce que chez le mâle. Cette
observation témoigne également de la surve-
nue précoce de ce dimorphisme (31). Chez le
mouton, un dimorphisme sexuel existe égale-
ment, mais il est inverse à celui observé chez
le primate. Le rétrocontrôle gonadique inhibi-
teur se lève à un âge beaucoup plus jeune
chez le mâle que chez la femelle. Chez la
femelle, l’administration prénatale de testo-
stérone entraîne, une “masculinisation” des
délais dans lesquels la puberté se déclenche et
suggère l’importance de la vie intra-utérine à
cet égard (32). Le métabolisme des stéroïdes
sexuels, en particulier l’aromatisation des
androgènes en estrogènes, peut également
jouer un rôle important dans les effets neu-
roendocriniens de ces stéroïdes. Récemment,
nous avons montré une potentialisation par
les estrogènes de la réponse GnRH induite
par le glutamate et cet effet serait de type non
génomique. Cela laisse entrevoir de nouvelles
possibilités dans les mécanismes des effets
des estrogènes sur la sécrétion de GnRH.
Cependant la signification physiologique de
ces observations reste à démontrer.
Signaux environnementaux
L’avancement séculaire de l’âge de la
ménarche s’est manifesté avec l’industrialisa-
tion de l’Europe et l’amélioration des condi-
tions hygiéno-diététiques, entre la moitié du
XIXesiècle et la moitié du XXesiècle. Les
premières règles sont apparues progressive-
ment plus tôt, passant de 17 à 13 ans.
L’influence de la nutrition sur l’axe hypo-
physo-gonadique est évidente, surtout chez la
fille. Ce facteur permet d’expliquer certaines
formes de retard de puberté. De nombreux
polluants environnementaux doués d’activité
estrogénique et/ou anti-androgénique pour-
raient avoir une influence significative sur
l’axe hypothalamo-hypophyso gonadique. En
Belgique, il a été observé qu’un quart des
patients vus pour puberté précoce sont des
fillettes étrangères arrivées en Belgique le
plus souvent dans le cadre d’une adoption ou,
parfois, avec leur famille d’origine (33). Cette
observation singulière a alors posé la question
du rôle éventuel d’une exposition préalable de
Mise au point

Act. Méd. Int. - Métabolismes - Hormones - Nutrition, Volume V, n° 3, mai-juin 2001
105
ces enfants dans leur pays d’origine aux xéno-
estrogènes sans que le mécanisme exact puisse
encore être élucidé.
S’il est admis que le rôle des signaux envi-
ronnementaux comme les phéromones est
assez évident chez l’animal, l’implication de
tels facteurs n’est pas à exclure chez l’hom-
me. Il a en effet été rapporté récemment que
l’exposition de la muqueuse olfactive à des
sécrétions apocrines recueillies en phase fol-
liculaire ou en période périovulatoire, rac-
courcissait ou allongeait respectivement la
phase folliculaire (34).
Conclusion
Le déclenchement de la puberté semble être
la résultante d’une série d’événements en cas-
cade aboutissant à l’accélération de la sécré-
tion pulsatile de GnRH sans que l’on puisse à
l’heure actuelle identifier les structures neu-
ronales et gliales qui vont exercer le contrôle
ultime et donc décisif sur la sécrétion de
GnRH. Les différences d’action et aussi de
sensibilité aux facteurs régulateurs d’origines
hypothalamique, périphérique et environne-
mentale pourraient expliquer la variabilité de
l’âge au déclenchement de la puberté.
L’émergence, par excès ou par défaut, d’un
des facteurs facilitateurs ou inhibiteurs est
susceptible d’entraîner une puberté anormale-
ment précoce ou tardive, selon le cas.
●
Références
1. Herman-Giddens ME, Slora EJ, Wasserman
RC et al. Secondary sexual characteristics and
menses in young girls seen in office practice : a
study from the Pediatric Research in Office
Settings Network. Pediatrics 1997 ; 99 : 505-12.
2. Mul D. Treatment of early puberty in adopted
and non-adopted children : when, why and how.
Auxological, psychological and ethical aspects
of growth promoting treatment around puberty.
Thesis Erasmus University Rotterdam 2000,
The Netherlands.
3. Knobil E. The neuroendocrine control of the
menstrual cycle. Recent Progress in Hormone
Research 1980 ; 36 :53-88.
4. Martinez de la Escalera G, Choi ALH,
Veiner RI. Generation and synchronization of
gonadotropin-releasing hormone (GnRH)
pulses : intrinsic properties of the GT1-1 GnRH
neuronal cell line. Proc Natl Acad Sci USA
1992 ; 89 :1852-5.
5.Terasawa E, Keen KL, Mogi K et al. Pulsatile
release of luteinizing hormone-releasing hor-
mone (LHRH) in cultured LHRH neurons deri-
ved from the embryonic olfactory placode of the
rhesus monkey. Endocrinology 1999 ; 140 :
1432-41.
6. Funabasshi T, Daikoku S, Shinohara K et al.
Pulsatile gonadotropin-releasing hormone
(GnRH) secretion is an inherent function of
GnRH neurons, as revealed by the culture of
medial olfactory placode obtained from
embryonic rats. Neuroendocrinology 2000 ;
71 :138-44.
7. Plant TM, Gay VL, Marshall GR et al.
Puberty in monkeys is triggered by chemical sti-
mulation of the hypothalamus. Proc Natl Acad
Sci USA 1999 ; 86 : 2506-10.
8. Bourguignon JP, Gérard A, Mathieu J et al.
Maturation of the hypothalamic control of pul-
satile gonadotropin-releasing hormone secre-
tion at onset of puberty : increased activation of
N-methyl-D-aspartate receptors. Endocrino-
logy 1999 ; 127 : 873-81.
9. Mitsushima D, Hei D, Terasawa E.
γ
-amino-
butyric acid is an inhibitory neurotransmitter
restricting the release of luteinizing hormone-
releasing hormone before the onset of puberty.
Proc Natl Acad Sci USA 1994 ; 91 : 395-9.
10.Yamanaka C, Lebrethon MC,Vandersmissen
E et al. Early prepubertal ontogeny of pulsatile
gonadotropin-releasing hormone secretion.
Inhibitory autofeedback control through prolyl
endopeptidase degradation of GnRH.
Endocrinology 1999 ; 140 : 4609-15.
11.Bourguignon JP, Gérard A, Purnelle G et al.
Duality of glutamatergic and gabaergic control
of pulsatile GnRH secretion by rat hypothala-
mic explants. II. Reduced NR2C- and GABAa-
receptor-mediated inhibition at initiation of
sexual maturation. J Neuroendocrinol 1997 ; 9 :
193-9.
12. Terasawa E, Fernandez DL. Neurobiolo-
gical mechanisms of the onset of puberty in pri-
mates. Endocrine Reviews 2001 ; 22 : 111-51.
13. Bourguignon JP, Jaeken J, Gérard A et al.
Amino acid neurotransmission and initiation of
puberty : evidence from nonketotic hyperglyci-
nemia in female infant and gonadotropin-relea-
sing hormone secretion by rat hypothalamic
explants. J Clin Endocrinol Metab 1997 ; 82 :
1899-903.
14. Ojeda SR, Ma YJ, Dziedzic B et al.
Astrocyte-neuron signaling and the onset of
female puberty. In : Bourguignon JP, Plant TM
(eds).The onset of puberty in perspective.
Elsevier Science BV, 2000 : 41-57.
15. Jung H, Carmel P, Schwartz MS et al. Some
hypothalamic hamartomas contain transfor-
ming growth factor alpha, a puberty-inducing
growth factor, but not luteinizing hormone
releasing-hormone neurons. J Clin Endocrinol
Metab 1999 ; 84 : 4695-701.
16. Wierman ME, Allen MP, Xiong X et al.
Developmental control of gonadotopin relea-
sing hormone (GnRH) gene expression : role of
POU homeodomain proteins. In : Bourguignon
JP, Plant TM (eds).The onset of puberty in pers-
pective. Elsevier Science BV, 2000 : 15-23.
17. Frisch RE, McArthur J. Menstrual cycles :
fastness as a determinant of minimum weight of
height necessary for the maintenance or onset.
Science 1994 ; 185 : 949-51.
18. Loucks AB, Heath EM. Dietary restriction
reduces luteinizing hormone (LH) pulse fre-
quency during waking hours and increases LH
pulse amplitude during sleep in young mens-
truating women. J Clin Endocrinol Metab
1994 ; 78 : 910-5.
19. Sisk CL, Bronson FH. Effects of food res-
triction and restoration on gonadotropin and
growth hormone secretion in immature male
rats. Biology of reproduction 1986 ; 35 : 554-
61.
20. Dubey AK, Cameron JL, Steiner RA et al.
Inhibition of gonadotropin secretion in castra-
ted male rhesus monkeys (Macaca mulatta)
induced by dietary restriction : analogy with the
prepubertal hiatus of gonadotropin release.
Endocrinology 1986 ; 118 : 518-25.
21. Zhang Y, Proenca R, Maffei M et al. Positional
cloning of the mouse obese gene and its human
homologue. Nature 1994 ; 372 : 425-32.
22. Chehab FF, Lim ME, Ronghue L et al.
Correction of the sterility defect in homozygous
obese female mice by treatment with the human
recombinant leptin. Nat Genet 1986 ; 12 : 318-20.
 6
6
1
/
6
100%