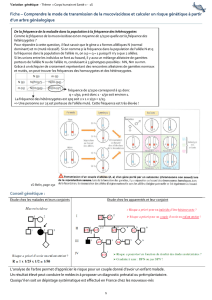
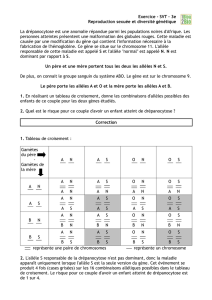
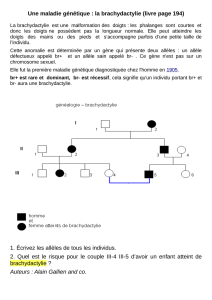
1 - Science Calender

• Début XXème siècle : U. Yule, W. Castle, G. Hardy and W. Weinberg, S. Wright,
R. Fisher, JBS Haldane : modèles mathématiques de la structure génétique des populations
• 1960-2000 : application des techniques de biochimie et génétique moléculaire pour étudier
les fréquences d’isoformes protéiques, d’allèles, aux niveaux protéique et ADN
Population P
Groupe d’individus partageant un
ensemble commun de gènes dans une
aire géographique, réellement ou
potentiellement interféconds
Evaluation des fréquences
alléliques, des différents
allèles de chaque gène (locus)
des fréquences génotypiques
dans la population et les variations
de ces fréquences d’une génération
À l’autre (n, n+1, n+2, n+n’)
Pr. A. CALENDER
Dr. C. BARDEL

HARDY GODFREY HAROLD (1877-1947)
Mathématicien anglais, né à Granleigh, dans le Surrey, et mort à Cambridge.
Godfrey Harold Hardy fit ses études au Trinity College de Cambridge, où il
enseigna de 1906 à 1919. En 1908, il découvre, en même temps que le
physicien W. Weinberg, mais indépendamment de lui, la loi de Hardy-
Weinberg, qui décrit l'équilibre génétique au sein d'une population et qui
aura une très grande importance pour l'étude des facteurs Rhésus dans les
groupes sanguins
Wilhelm WEINBERG (1862-1937)
Gynécologue obstétricien et physicien, il publia en 1908, de
manière indépendante les principes de base de la Loi. Son travail
ne fut reconnu que 35 ans après et associé au travail de
HARDY. Probablement fut il le premier, mais son article rédigé en
allemand fit que cette reconnaissance a été tardive, associée
au fait qu’il fut le premier également à reconnaître les biais de
la loi, reconnus aujourd’hui comme écarts à la Loi de HW
Pr. A. CALENDER – 2010-2011

Les génotypes sont ils distribués au hasard dans l’espace et dans le temps ?
Existe-t-il une structuration de la variation ?
Quels sont les processus qui affectent le pool de gènes de la population ?
Ces processus induisent-ils une divergence entre les populations ?
F(a)
Temps (générations)
0,5


 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
1
/
68
100%