CHAPITRE 2 : Équilibre de Hardy Weinberg - FSA

Génétique des populations SV5 chapitre 2 Page 15
CHAPITRE 2 : Équilibre de Hardy Weinberg
Dans un cas simple appelé p o p u l a t i o n t h é o r i q u e i d é a l e , qui se
définit par les caractéristiques suivantes :
1) population d'organismes d i p l o ï d e s à r e p r o d u c t i o n s e x u é e et à
g é n é r a t i o n s n o n c h e v a u c h a n t e s (aucun croisement entre individus
de générations suivantes)
2) population d'e f f e c t i f i n f i n i où les croisements sont entièrement
a l é a t o i r e s
3) p o p u l a t i o n c l o s e génétiquement (absence de flux migratoires)
4) tous les individus, quel que soit leur génotype, ont la même capacité à se
reproduire et à engendrer une descendance viable =
absence de sélection
5) A b s e n c e d e m u t a t i o n et de distorsion de ségrégation méïotique
(un individu
Aa
produira toujours 50% de gamètes
A
et 50% de gamètes
a
).
Parmi toutes ces caractéristiques, le croisement au hasard des individus,
appelé système de r e p r o d u c t i o n p a n m i c t i q u e , est l'hypothèse la
plus importante. Cette hypothèse suppose que les individus ne choisissent pas
leur partenaire sexuel ni en fonction de leur génotype, ni en fonction de leur
phénotype = p a n m i x i e et que la rencontre des gamètes se fait au hasard =
p a n g a m i e .
A - CONSTITUTION GENETIQUE DES POPULATIONS
1- Définitions :
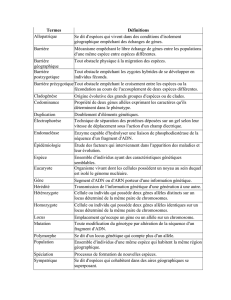
Pour définir la constitution génétique d'une population plusieurs niveaux,
phénotypique, génotypique et génique sont utilisés. Ils sont décrits par une loi de
probabilité. Ainsi pour une variable donnée (le caractère: couleur des yeux) on
précisera les différents états possibles (bleu, vert, marron ...) et les probabilités
avec lesquelles ces états se rencontrent dans la population étudiée (0,10 ; 0,20 ;
0,60 ...). Ce premier descripteur est la constitution phénotypique de la population.
2- Exemples d'Estimation de constitutions génétiques
2.1 Cas de gènes autosomiques
Soit une population de N individus diploïdes à reproduction sexuée biparentale,
étudiée pour un caractère à déterminisme monogénique. On y observe trois classes

Génétique des populations SV5 chapitre 2 Page 16
génotypiques d'effectifs respectifs (A1A1) n1, (A1A2) n2, (A2A2) n3, avec n1 + n2 +
n3 = N. Le déterminisme génétique étant connu, un couple d'allèles autosomiques A1,
A2 codominants, la constitution génotypique de la population correspond à sa
constitution phénotypique. Les fréquences génotypiques sont alors exactement
définies par les rapports :
freq (A1A1) = f1 = D = n1/N freq (A1A2) = f2 = H = n2/N
freq (A2A2) = f3 = R = n3/N
p = (2n1 + n2)/2N = D + 1/2 H
q = (2n3 + n2)/2N = R + 1/2 H.
2.2 Cas de gènes liés au sexe (ou de gènes d'un système haplo-diploïde) :
Soit A1 et A2 des allèles situés sur l'hétérochromosome commun aux deux sexes.
Dans une population où l'on dénombre N1 individus de sexe homogamétique et N2 de
sexe hétérogamétique, le nombre de gènes concernés est de 2N1 + N2. Notons n1,
n2, n3, les effectifs des génotypes A1A1, A1A2, A2A2 chez les femelles, et m1 et
m2 ceux des génotypes A1 et A2 chez les mâles. En considérant chaque groupe
sexuel comme un échantillon indépendant, on peut estimer la fréquence de l'allèle A1
pour chacun des sexes et pour l'ensemble de la population :
p= (2n1 + n2)/2N1 p’ = m1/ N2 P = (2n1 + n2 + m1)/(2N1 + N2)
(femelles) (mâles) (l’ensemble de la population)
B- DEFINITION DU MODELE DE HARDY-WEINBERG
1 - L'Hypothèse panmictique :
1.1 CAS DE GENES AUTOSOMIQUES
1. 1. 1 RENCONTRE DES GAMETES
Lorsque la rencontre des gamètes se fait au hasard, on dit qu'il y a pangamie;
lorsque la rencontre des individus se fait au hasard, ont dit qu'il y a panmixie.
Dans l'hypothèse panmictique, chaque nouveau zygote formé résulte de deux tirages
au sort indépendants, l'un dans l'urne des ovules, l'autre dans l'urne des
spermatozoïdes. Ces urnes sont respectivement composées de l'ensemble des ovules
matures fournis par les femelles de la population au moment du tirage et de
l'ensemble des spermatozoïdes matures fournis par les mâles de la population au
moment du tirage. Les probabilités de chaque nouveau zygote sont donc :
A1A1 = p2 A1A2 = 2pq A2A2 = q2.
On retrouve la relation :
(p +q) 2 = p2+ 2pq + q2 = 1,
1.1.2 RENCONTRE BASE SUR LES CROISEMENTS DES INDIVIDUS

Génétique des populations SV5 chapitre 2 Page 17
Considérons le cas d’un locus avec deux allèles et une population infinie où les
fréquences des trois génotypes ont des valeurs quelconques à la génération n.
Génotypes et fréquences des mâles
A1A1 A1A2 A2A2
D H R
________________________________
Génotypes A1A1 D D2 DH DR
et fréquences A1A2 H DH H2 HR
des femelles A2A2 R DR HR R2
A1A1 A1A2 A2A2
A1A1 x A1A1 D2 1 - -
A1A1 x A1A2 2DH 1/2 1/2 -
A1A1 x A2A2 2DR - 1 -
A1A2 x A1A2 H2 1/4 1/2 1/4
A1A2 x A2A2 2RH - 1/2 1/2
A2A2 x A2A2 R 2 - - 1
total 1 D1 H1 R1
D1 = D2 + 1/2(2DH) +1/4(H2) = p2
H1= 1/2(2DH) + (2DR) + 1/2H2 +1/2(2RH) = 2(D+1/2H)(R+1/2H) = 2pq
R1 = R2 +1/2(2RH) + 1/4 H2 = q2
La structure génotypique restera identique au cours des générations ultérieurs
puisque les proportions D, H et R ne dépendent que de p et q .
2. Cas d'un locus situé sur un hétérochromosome
(et gènes d'un système haplo-diploïde)
Lorsqu'un gène est situé sur un hétérochromosome, le sexe homogamétique
possède deux locus et le sexe hétérogamétique un seul.
Si les deux sexes sont en nombre égal dans la population, 2/3 des allèles liés au sexe
seront portés par le sexe homogamétique(p) et 1/3 par l'autre sexe(p’). Pour un taux
sexuel différent, il suffit de pondérer les proportions par les effectifs de chaque
sexe.
P=2/3p + 1/3p’
2.1 CONSTITUTIONS GENETIQUES A L'ETAT D'EQUILIBRE
femelles mâles
A1 A1 A1 A2 A2 A2 A1 A2
p2 2 p q q2 p’ q’
La fréquence de l'allèle A1 est égale à p=p’ dans chaque sexe, elle restera inchangée
dans les deux sexes après une génération en reproduction panmictique. Le même

Génétique des populations SV5 chapitre 2 Page 18
raisonnement est vrai quel que soit le numéro d'ordre de la génération. Nous sommes
en présence d'un état d'équilibre.
2.2 L'ATTEINTE DE L'ETAT D'EQUILIBRE
Même si la fréquence allélique n'est pas identique dans les deux sexes, il est évident
que, sous les hypothèses de la loi de Hardy-Weinberg, la fréquence allélique pour
l'ensemble de la population ne variera pas puisque aucun facteur évolutif n'agira. Par
contre, nous allons montrer que la distribution des fréquences alléliques, dans
chaque sexe, varie en oscillant, tout en se rapprochant de la position d'équilibre.
p’ n = p n-1 p n = 1/2 pn-1+ 1/2 p’ n-1 En = p n-p’ n Eo = po-p’o En=(-1/2)nEo
On déduit alors les relations :
Lorsque n augmente, (-1/2)n tend vers zéro et la fréquence allélique A1 des femelles
tend vers p=pn=p’n=(2p0+p’0)/3.
A titre d'exemple considérons le cas extrême d'une population de base dans
laquelle toutes les femelles sont A2A2 et tous les mâles A1Y. Soit 0 la fréquence de
l'allèle A1 chez les femelles et 1 chez les mâles.
C - SYSTEMES MULTI-LOCUS
1- Etude d'un système à deux locus autosomiques :
1.1 Constitutions gamétique et génotypique à l'équilibre
Soient deux locus différents A et B pouvant être occupés le premier par les
allèles A ou a et le second par les allèles B ou b. Il n'est pas nécessaire de dissocier
le cas de locus situés sur un même chromosome (physiquement liés) de celui de locus
situés sur des chromosomes différents, étant donné que la probabilité (c) de
recombinaison entre les deux locus n'intervient pas sur la constitution génétique à
l'équilibre, comme nous le verrons ultérieurement. Dans le cas de locus physiquement
liés, (c) est comprise entre 0 et 1/2 si ces locus sont suffisamment proches
(génétiquement liés) et égale à 1/2 si les locus sont très éloignés (génétiquement
indépendants). (c) est également égale à 1/2 si les locus sont situés sur des
chromosomes différents (physiquement indépendants).
(pA + pa)2 et ( pB + pb)2
Pour les deux locus pris simultanément, la constitution génotypique globale, à
l'équilibre, sera donnée par le développement du produit de ces binômes soit :
2po+ p’o po - p’o
p’n = ------------------- + (-1 /2)n -1 ------------
3 3
2po+ p’o po - p’o
pn = --------------------- + (-1 /2)n ------------
3 3

Génétique des populations SV5 chapitre 2 Page 19
[(pA + pa)2 (pB +pb)2] .
Il existe quatre types de gamètes possibles : AB, Ab, aB et ab, de fréquences
respectives pApB, pApb, papB et papb. On donne souvent le nom d'haplotype à ces
combinaisons gamétiques: par exemple haplotype AB. Si les gènes considérés sont
associés au hasard dans les gamètes, la fréquence de chaque haplotype sera
simplement égale au produit des fréquences des allèles respectifs qui le constitue.
Dans ce cas la population est à l'état d'équilibre de Hardy-Weinberg et on dit à
l’équilibre d'association gamétique ou simplement équilibre d'association. On emploie
parfois le terme d'équilibre de liaison (
linkage equilibrium
). Cette appellation n'est
cependant pas très appropriée dans la mesure où elle laisse croire qu'il s'agit de
gènes physiquement liés. En fait le raisonnement est valable pour tous les locus. Les
états d'équilibre des différentes constitutions génétiques seront identiques, que les
locus soient physiquement liés ou non. Comme nous le démontrerons, seule la vitesse
d'atteinte de l'état d'équilibre dépend du pourcentage de recombinaison.
Si à une génération donnée l'association aléatoire des allèles n'est pas réalisée
lors de la formation des gamètes, certains haplotypes présenteront des fréquences
supérieures, ou inférieures, à celles attendues à l'équilibre. La population n'est donc
pas à l'état d'équilibre. Pour l'haplotype AB on peut alors écrire :
AB = pApB + D,
où D est une valeur relative qui représente le déséquilibre d'association gamétique
(dit aussi, mais de façon moins correcte, déséquilibre de liaison ou "
linkage
disequilibrium
"). Les fréquences des autres haplotypes seront modifiées avec la
même valeur absolue. En effet, si la fréquence d'association des allèles est
modifiée, les fréquences alléliques, elles, ne le sont pas, car la population est
toujours régie par le principe de Hardy-Weinberg. La somme des fréquences des
haplotypes AB et Ab par exemple, qui regroupent tous les allèles A, doit être égale à
pA. Ceci implique que la fréquence des haplotypes Ab soit :
Ab = pApb - D.
Dans ces conditions D est différent de zéro et la population est effectivement
en déséquilibre d'association gamétique. Les fréquences gamétiques réelles,
fonctions des fréquences à l'équilibre et de D (Lewontin et Kojima, 1960, Evolution
14:458), s'écrivent :
AB = pApB + D et D=AB - pApB
Ab = pApb - D
aB = papB - D
ab = papb + D,
On en déduit alors : D = AB.ab – aB.Ab.
2 - L'approche de l'état d'équilibre
L'atteinte de l'état d'équilibre est fonction du degré de recombinaison entre les
locus (génétiquement indépendants: c=0.5, ou non indépendants: c < 0,5).
 6
6
1
/
6
100%