Virus et Muqueuses

Virus et Réponses Immunitaires Muqueuses
Jennifer Richardson
UMR 1161 Virologie INRA AFSSA ENVA

Les réponses immunitaires anti-virales:
Pourquoi s’y intéresser ?
Biologie appliquée :
Mise au point des vaccins anti-viraux
Développement des vecteurs viraux pour le transfert de gène
(codant un immunogène/ gène thérapeutique)
Biologie fondamentale :
Comprendre la co-évolution des espèces cellulaires & virales

Virus et Muqueuses
Introduction
Virus et Barrière Muqueuse
Barrières Extrinsèques
Barrières Intrinsèques
Voies d’entrée in vivo
Virus et Immunité Innée
Immunité innée et acquise
Spécificités des infections virales
Phase d’induction
Réponses impliquant les interférons
Cellules NK
Virus et Immunité Acquise
Organisation du système immunitaire muqueux
Phases d’induction et effectrice
Anticorps de sécrétion
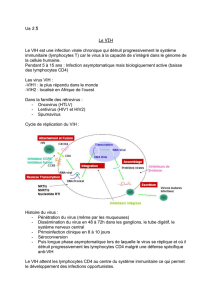
Virus de l’Immunodéficience Humaine

Virus et Muqueuses
Introduction : Virus
Les Virus sont des micro-organismes acellulaires & des parasites intracellulaires
obligatoires.
Adapté de
Bomsel M. & Alfsen A. 2003.
Nature Rev. Mol. Cell Biol. 4: 57-68.
Virus enveloppé
(acides nucléiques + protéines
+ membrane)
Virus nu
(acides nucléiques
+ protéines
Génome ADN ou ARN Protéines
capsidales Membrane (Péplos)
(lipides & protéines)
Glycoprotéine d’enveloppe
virale
ou
-

Virus et Muqueuses
La classification des virus repose sur leur structure.
Introduction : Virosphere
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
1
/
80
100%