Chapitre 4 : Un regard sur l`évolution de l`Homme

CHAPITRE 4 : UN REGARD SUR
L'ÉVOLUTION DE L'HOMME

Introduction
Comme toute autre espèce, l’Homme actuel, Homo sapiens, résulte d’une
évolution. Son histoire fait partie de celle du genre humain (Homo), qui
a comporté d’autres espèces aujourd’hui disparues, et s’inscrit d’une façon
plus générale dans celle des primates.
1. La diversité actuelle et passée des primates
A. La diversité des primates
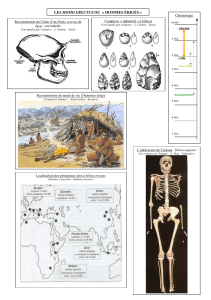
Problème : Quels sont les critères d'appartenance au genre humain ?

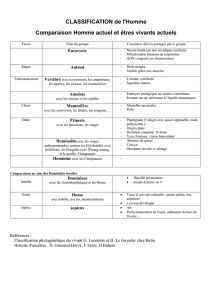
Les Primates
Principales caractéristiques
51 genres, 168 espèces
•Pouce et gros orteil opposables aux
autres doigts (sauf chez les humains pour
le gros orteil), permet la préhension.
•Ongles plats à la place des griffes (sauf
quelques petites espèces d’Amérique du
Sud).
•Yeux rapprochés sur le devant de la
face ( permet la vision binoculaire).
•L' appendice nasal est un nez et non
une truffe (Museau réduit).
•Cerveau développé (capacité au moins
égale à 350 à 400 cm3).
•D'autres caractéristiques : organisation
sociale et familiale, communication
importante, tendance à se redresser…

Des
Primates…
dont l'Homme fait
partie.

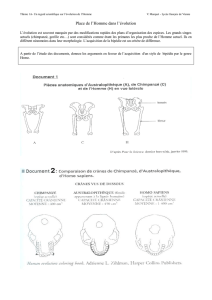
1. La diversité actuelle et passée des primates
B. L'établissement d'une phylogénie
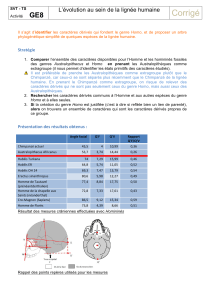
Correction du TP 8
Avec le logiciel Phylogène, on construit la matrice suivante :
Le Toupaïe n'appartenant pas aux primates, il est choisit comme extra
groupe. Cela signifie que tous ses caractères sont considérés comme des
caractères ancestraux et donc la présence d'un caractère différent est
considéré comme un caractère dérivé, c'est une innovation évolutive
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
1
/
46
100%