Web 2 Fiche 2

1
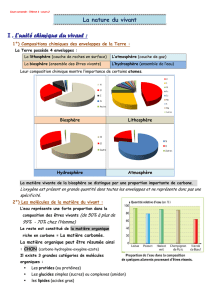
Web 2 Fiche 2.1
CYCLE DE KREBS
G. Ona-Nguema, C. Roose-Amsaleg et P. Bauda
Le cycle de Krebs permet d’oxyder complètement les molécules d’acétate (issues de l’oxydation
du pyruvate) en dioxyde de carbone et donc de produire une grande quantité de cofacteurs
réduits (NADH,H+ et FADH2) qui vont fournir les électrons à la chaîne respiratoire.
L’oxaloacétate (OAA) est une molécule très importante, dont le déficit dans la cellule engendre
un dysfonctionnement du cycle de Krebs. Notons que C4, C5 et C6 désignent le nombre de
carbone des molécules impliquées dans le cycle.

2

3
Web 2 Fiche 2.2
GLYCOLYSE OU VOIE D’EMBDEN-MEYERHOF
G. Ona-Nguema, C. Roose-Amasaleg et P. Bauda
La glycolyse est une chaîne de 10 réactions enzymatiques : les réactions 1 à 4 font intervenir les
molécules à 6 carbones (partie I), alors que les réactions 5 à 10 impliquent les composés à 3
carbones (partie II). Les enzymes catalysant ces réactions sont : (1) hexokinase, (2)
phosphoglucose isomérase, (3) phosphofructokinase, (4) aldolase, (5) triose-phosphate
isomérase, (6) glycéraldéhyde-3-phosphate déshydrogénase, (7) phosphoglycérate kinase, (8)
phosphoglycérate mutase, (9) énolase, (10) pyruvate kinase.
℗ désigne le groupement phosphate.

4

5
Web 2 Fiche 2.3
VOIE DES PENTOSES PHOSPHATES
G. Ona-Nguema, C. Roose-Amsaleg et P. Bauda
La voie des pentoses phosphates peut être divisée en deux grandes parties : (i) la branche
oxydative, composée des réactions 1 à 3 (en noir), au cours de laquelle il y a oxydation d’une
molécule de glucose-6-phosphate avec libération du dioxyde de carbone, d’un pentose
phosphate et de deux NADPH,H+ ; et (ii) la branche non-oxydative représentée par des réactions
4 à 11 (en orange). Les enzymes catalysant ces réactions sont : (1) glucose-6-phosphate
déshydrogénase, (2) phosphogluconolactonase, (3) 6-phosphogluconate déshydrogénase, (4)
pentose-phosphate isomérase, enzyme qui permet une isomérisation réversible du cétose
(ribulose-5-phosphate) en aldose (ribose-5-phosphate), (5) pentose-phosphate épimérase,
enzyme permettant une épimérisation réversible au niveau du carbone 3 des cétoses (ribulose-5-
phosphate en xylulose-5-phosphate), (6) transcétolase, enzyme qui échange deux carbones entre
un cétose (donneur) et un aldose (accepteur) (7) transaldolase, enzyme qui échange trois
carbones entre un cétose (donneur) et un aldose (accepteur), (8) triose-phosphate isomérase, (9)
aldolase, (10) phosphofructokinase ou fructose-bisphosphatase, (11) phosphoglucose isomérase
(Adapté de Kruger et von Schaewen, 2003).
℗ désigne le groupement phosphate.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
1
/
21
100%