Chapitre 1 – Méiose, fécondation et diversité génétique

Chapitre 1 – Méiose, fécondation et diversité génétique
Introduction
Une espèce est un ensemble d’individus qui se ressemblent morphologiquement, qui sont
interféconds et dont la descendance est fertile. Au niveau génétique, ils ont le même caryotype (sauf
différence entre les sexes) (…… chromosomes = …… paires dans l'espèce humaine) et possèdent
les mêmes gènes (25000 dans l'espèce humaine). Comment expliquer le maintien au fil des
générations des caractéristiques génétiques d'une espèce (son caryotype notamment) ?
Au sein d’une espèce, tous les individus sont génétiquement différents. C'est également vrai pour les
enfants d'un même couple (sauf cas des vrais jumeaux).
Comment expliquer cette diversité génétique ?
I. Les phénomènes chromosomiques de la méiose
Le nombre de chromosomes passe de ……… dans la plupart des
cellules de l'organisme à ……… dans les cellules sexuelles, pour
revenir à ……… après la fécondation.
Par quels mécanismes ?
Chapitre 1 - Méiose, fécondation et diversité génétique 1 / 18
……
……
……
……
……
……
……
……

Activité 1 – Les événements de la méiose
1. Combien de divisions cellulaires successives distingue-t-on au cours de la méiose ? ………………
2. A l’aide de la vidéo, remplir le tableau suivant des différentes étapes de la méiose :
Phase Nombre de
cellules
Chromosomes : combien dans chaque cellule (2×n ou 1×n, où
n est le nombre de paires de chromosomes) ?
à combien de chromatides chacun ?
4. Situer par une flèche le moment de la réduction chromatique (passage diploïde → haploïde).
5. Construire un graphique indiquant l’évolution de la quantité d’ADN en fonction du temps dans
les cellules au cours de la méiose. On note Q la quantité d’ADN au début de l'interphase précédant
la méiose.
Chapitre 1 - Méiose, fécondation et diversité génétique 2 / 18
Temps
Quantité d'ADN
par cellule
Q

Chapitre 6 - Méiose, fécondation et diversité génétique 3 / 18

Bilan du I :
L'observation du caryotype d'une cellule somatique (peau, muscles, etc) montre que les chromosomes
peuvent être regroupés par paires de chromosomes de même type (même taille, même position du
centromère, même disposition des bandes de coloration) : les chromosomes d'une même paire sont dits
homologues. Ils portent la même série de gènes (mais pour chaque gène, pas nécessairement le même
allèle). Une cellule somatique est qualifiée de diploïde. Son caryotype est noté 2n (n étant le nombre de
paires).
Dans les gamètes, spermatozoïdes et ovules, on ne compte en revanche qu'un seul exemplaire de
chaque type de chromosome : ces cellules sont haploïdes, comportant n chromosomes
(un chromosome de chacune des n paires) .
La méiose est une succession de deux divisions cellulaires particulières qui permet de produire des
gamètes haploïdes à partir d'une cellule germinale diploïde. Chez les animaux, elle se déroule dans les
testicules et les ovaires.
Durant l'interphase qui précède la méiose, il y a réplication de la molécule d'ADN de chaque chromosome. Au
début de la méiose, les chromosomes apparaissent donc dédoublés en deux chromatides identiques.
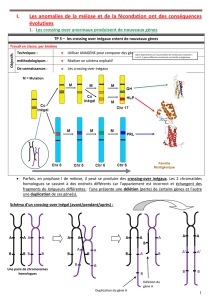
On considère une cellule (animale ici) contenant 2 paires de chromosomes. Sur le schéma de la
page suivante, on dessinera la paire n°2 sensiblement plus grande. Chaque paire comportera un
chromosome rouge et un bleu. Sur la paire n°1, on placera un seul gène A, à l’état hétérozygote. Sur
la paire n°2, on placera 3 gènes :
- le gène B à l’état hétérozygote;
- le gène D à l'état hétérozygote.
Les quatre cellules produites par la méiose contiennent-elles la même information génétique ?
……………………………………………………………………………………………………………………
……………………………………………………………………………………………………………………
Chapitre 1 - Méiose, fécondation et diversité génétique 4 / 18
Schéma pour
information

Prophase I
Point de départ : une cellule germinale à
2n chromosomes (= diploïde).
Disparition de l'enveloppe nucléaire.
Condensation de l'ADN des chromosomes
puis les chromosomes homologues
s'apparient étroitement. (Deux
homologues forment un bivalent, aussi
appelé tétrade car 2×2=4 chromatides.)
A ce moment, les homologues échangent des fragments de
chromatide. A la fin de la prophase, les chromosomes homologues sont
un peu moins étroitement accolés et laissent voir des chiasmas, traces
d'un échange de segment de chromatide.
Métaphase I
Les tétrades se disposent à l'équateur
de la cellule, les centromères de chaque
chromosome se plaçant de part et d'autre
du plan équatorial.
Anaphase I
Les chromosomes homologues se
séparent et migrent chacun vers un
pôle de la cellule.
Chaque chromosome conserve ses
deux chromatides.
Télophase I
La cellule se coupe en deux.
Chaque cellule contient alors n
chromosomes , un de chaque paire : elle
est haploïde. Le nombre de
chromosomes a été divisé par deux :
division réductionnelle.
Mais chaque chromosome a encore ses
deux chromatides.
Dans ses manifestations
chromosomiques, la deuxième
division est semblable à une
mitose. Elle ne change pas le
nombre de chromosomes : division
équationnelle.
Prophase II
Métaphase II
Les chromosomes se placent de telle
façon que leur centromère soit dans
le plan équatorial.
Anaphase II
Les deux chromatides de chaque
chromosome se séparent et
chacune migre vers l'un des pôles.
Télophase II
L'ADN des chromosomes se
décondense. L'enveloppe nucléaire
se reforme.
Chaque cellule se coupe en deux.
Les cellules issues de la méiose
possèdent un seul chromosome de
chaque paire (n chromosomes),
elles sont donc haploïdes .
Chaque chromosome n'est plus
formé que d'une seule chromatide.
Chapitre 6 - Méiose, fécondation et diversité génétique 5 / 18
Première division de méiose (division réductionnelle)
Deuxième division de méiose (division équationnelle, semblable à une mitose)
 6
7
8
9
10
11
12
13
14
15
16
17
18
6
7
8
9
10
11
12
13
14
15
16
17
18
1
/
18
100%