le muscle

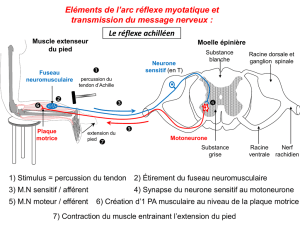
1
LE MUSCLE
Le handball est une activité physique qui présente la particularité de concerner l’ensemble des
muscles et des chaînes musculaire de l’organisme, mais aussi tous les types de fibres.
1 STRUCTURE MUSCULAIRE
1.1-Contenu cellulaire
Les cellules musculaires sont des cellules allongées contenant plusieurs noyaux. La longueur d’une
cellule musculaire est très variable d’un muscle à l’autre mais certaines peuvent avoir la longueur du
muscle.
Le muscle contient :
+ Un sarcolemme (fine membrane élastique qui limite à l’extérieur la fibre musculaire striée.
Myofibrilles
Z Z Z
Sarcomère
Fibre musculaire
+ Des myofibrilles, orientées parallèlement les unes aux autres dans l’axe du muscle. Ces
myofibrilles sont groupées en sarcomères, délimitées par la strie Z.
Mitochondries
Z
Tubules Citernes
Un cytoplasme, ou sarcoplasme, contenant, outre les myofibrilles, des inclusions de
glycogène et de lipides, la myoglobine et les organites cellulaires comme le réticulum (lieu de
synthèse des protéines) et les mitochondries. On trouve également dans le sarcoplasme des citernes
transversales séparées du système tubulaire longitudinal, mais reliées entre elles par un système appelé
triade. Ces citernes sont destinées à contenir le Ca++ indispensable pour la contraction musculaire.

2
1.2-Protéines musculaires
+ Myofilaments
Les filaments comprennent les filaments de myosine, épais, libres au milieu du sarcomère, et
les filaments de myosine, plus fins, fixés à la membrane du sarcomère (strie Z)
Actine
Myosine
Z Sarcomère
La myosine présente un corps et une tête, grossièrement semblable à un club de golf. La tête
représente environ le 1/6 ème de la longueur totale. La partie céphalique ou méromyosine lourde est le
siège de l’activité enzymatique de la myosine (capacité d’hydrolyser l’ATP). Le segment distal, ou
queue, présente une affinité pour les molécules de myosine voisines (méromyosine légère).
Filament de myosine
Méromyosine
légère
La jonction entre la « tête » et la « queue » est mobile, donnant ainsi à la tête la capacité de s’orienter
Lors de la contraction musculaire la tête se fixe au ligament d’actine, puis opère sa rotation (environ
45°)
L’actine présente la forme d’une double hélice formée de deux chaînes monomériques
+ Protéines régulatrices de la contraction musculaire
Outre l’actine et la myosine, la contraction musculaire nécessite la présence de deux autres protéines,
la troponine et la tropomyosine.
La tropomyosine est formée d’un filament fin qui s’enroule autour de l’actine dans le sillon
formé par la double hélice. Un filament de tropomyosine concerne 7 monomères d’actine.
La troponine est une protéine globulaire qui chevauche la molécule de tropomyosine vers
l’une de ses extrémités. Cette protéine présente un site de fixation pour le Ca++.

3
2 CONTRACTION MUSCULAIRE
La contraction musculaire est le résultat :
De la fixation du Ca++ sur la troponine
De l’activation de la tropomyosine
De la fixation de la myosine sur l’actine
De l’hydrolyse de l’ATP, responsable de la mobilité de la tête de myosine
+ Au repos
Le calcium est enfermé dans les citernes cytoplasmiques
La troponine inhibe, via la tropomyosine, toute liaison de l’actine et de la myosine
De l’ATP est fixé sur la myosine
Les deux filaments sont libres et dans l’incapacité d’établir des liaisons.
+ Excitation
L’excitation du moto neurone alpha est responsable d’une activation de la plaque motrice.
Cette activation libère le Ca++ contenu dans le système cisternal.
Le calcium se lie à la troponine, libérant ainsi son inhibition vis à vis de la tropomyosine.
Ca ++
Mg++ Ca++
Troponine
Tropomyosine
Inhibition Libération du site
L’inhibition est levée
Cette activation Troponine/tropomyosine modifie le rapport Actine/tropomyosine (le ligament
de tropomyosine se déplace vers l’axe de la rainure formée par les deux brins de l’actine).
Les monomères d’actine sont alors libérés de leur inhibition, les têtes de myosine peuvent se
fixer sur l’actine.
+ Contraction
L’ATP qui était déjà fixé sur les molécules de myosine sous une forme dite « énergétisée »
(Mg+-ATP) provoque la brusque rotation des têtes de la myosine. L’hydrolyse de l’ATP fournit
l’énergie nécessaire à la contraction. Le filament d’actine glisse le long de celui de myosine,
rapprochant ainsi deux stries Z. Une plaque motrice concernant plusieurs sarcomères et un neurone
plusieurs plaques motrices, la contraction peut se développer dans l’ensemble du muscle.
Relaxation Contraction
Plaques motrices Z Z Z Z

4
+ Relaxation
L’ATP présent dans la cellule se fixe sur les têtes de myosine, séparant ainsi l’actine de la
myosine. Le calcium est repompé de façon active dans les citernes cytoplasmiques grâce à une
protéine, la calciquestrine. Ce phénomène est consommateur d’énergie, un ATP pour 2 Ca++.
La diminution de la calcicytie (calcium cytoplasmique) redonne à la troponine sa configuration
initiale, inhibitrice de la tropomyosine.
3 DIFFERENTS TYPES DE FIBRES
3.1 FIBRES LENTES ET RAPIDES
L’étude microscopique et physiologique permet de différencier plusieurs types de fibres. Ces
différences portent sur leur mode de fonctionnement énergétique, mais en aucun cas sur le mécanisme
de la contraction musculaire lui-même. Il faut se garder, pour schématiser ces différences, de
caractériser de façon trop systématique le mode de fonctionnement de ces cellules qui , lors de la
contraction musculaire interviendront toutes en synergie.
Deux grands types peuvent être distingués
Les cellules dites lentes, capable de contractions soutenues et d’exercices prolongés (type I)
Les cellules dites rapides, essentiellement recrutées lors des exercices en vitesse/intensité.
Ces deux types cellulaires présentent les mêmes voies métaboliques énergétiques, mais dans des
proportions différentes.
+ Les cellules lentes, grandes consommatrices d’acides gras, présentent de nombreuses
mitochondries, siège des processus oxydatifs. Elles sont aussi dites « rouges » car le transport de
grandes quantités d’oxygène des érythrocytes vers les mitochondries requière une forte concentration
de transporteur intracellulaire, la myoglobine (protéine proche de l’hémoglobine mais monomérique).
+ Les cellules rapides sont richement pourvues en enzymes ne nécessitant pas la présence
d’oxygène (voie glycolytique, système créatine/phosphate). Ces cellules présentent néanmoins des
mitochondries mais en quantité moindre. Leur moins grande concentration en myoglobine en fait des
cellules « blanches ». Ce type cellulaire peut être lui-même dissocié en II a (potentiel oxydatif élevé)
et II b très faiblement pourvu en mitochondries. On a également défini un type II c de cellules
intermédiaires, peu différenciées qui ne représentent chez l’homme que 1% des cellules musculaires
squelettiques.
Ces cellules sont également caractérisées par la taille du motoneurone et par la fréquence des
impulsions nerveuses. Les cellules lentes présentent un petit motoneurone et des fréquences de
stimulations basses (10 Hz), les cellules rapides, à l’inverse répondent à un grand motoneurone
générateur de fréquence élevée (100 Hz).
La proportion de fibres lentes et rapides est spécifique de l’espèce concernée. De plus au sein d’une
même espèce cette proportion est fixée génétiquement. Il existe donc spontanément dès la naissance
des sujets plus ou moins « doués » pour des exercices requérant de plus ou moins grandes capacités
aérobies. Dans l’espèce humaine il n’existe pas de différence liée au sexe.

5
La proportion de fibres lentes dans un muscle peut varier de 10 à 95%. Ainsi, dans le triceps sural, les
jumeaux contiennent environ 50% de cellules lentes pour près de 70% dans le muscle soléaire.
I II a II b
Motoneurone Petit Grand Grand
Fréquence Stimulation Faible Elevée Elevée
Densité capillaire Grande Moyenne Faible
Teneur en myoglobine Elevée Moyenne Faible
Teneur en glycogène Identique
Enzymes glycolytiques Faible Moyen Forte
Enzymes oxydatifs Elevé Moyen Faible
Nombre de mitochondries Elevé Moyen Faible
De la même façon, la synthèse des enzymes oxydatifs mitochondriaux dépend du DNA mitochondrial.
Or, lors de la fécondation, seules les mitochondries de la mère sont utilisées. Ce phénomène explique
pourquoi seules les capacités oxydatives de la mère peuvent être prises en compte lors de l’évaluation
d’un jeune athlète. Certaines populations présentent au niveau de leur génome mitochondrial des
capacités oxydatives particulièrement développées susceptibles d’en faire des athlètes particulièrement
redoutables dans les courses de 5000,
10 000 m ou plus.
3.2-EFFETS DE LA CROISSANCE
Le nombre global de cellules musculaires est fixé génétiquement. La croissance du muscle
répond donc à un réarrangement intrafibrillaire et non pas à une multiplication cellulaire.
+ L’allongement musculaire pendant l’enfance est dû à une augmentation du nombre
des sarcomères aux extrémités des fibres. Il n’existe pas d’allongement des sarcomères.
+ L’accroissement de la section musculaire pendant la période de croissance
correspond à une multiplication longitudinale des myofibrilles musculaires.
3.3-EFFETS DE L’ENTRAÎNEMENT
L’entraînement intervient de manière très significative sur la cellule musculaire. Ses effets
peuvent être perçus :
Dans la proportion des différents types de fibres II contenues dans le muscle.
La quantité de protéines contractiles contenue dans une fibre
La concentration des enzymes participant aux différentes filières.
Le nombre de mitochondries
Les réserves glycogéniques.
En ce qui concerne le type cellulaire il est possible de passer assez facilement du type II a au
type II b et inversement. Les entraînements en intensité pure (départ de sprint, sauts, impulsions, tirs...)
 6
7
8
9
6
7
8
9
1
/
9
100%