UV_101_BASES_PHYSIO_CONTRACT_MUSC

1
BASES DE PHYSIOLOGIE
UV 101
PHYSIOLOGIE
NEURO-MUSCULAIRE
P. PILARDEAU
I - LE MUSCLE

2
STRUCTURE MUSCULAIRE
1-Contenu cellulaire
Les cellules musculaires sont des cellules allongées contenant plusieurs noyaux. La longueur
d’une cellule musculaire est très variable d’un muscle à l’autre mais certaines peuvent avoir la
longueur du muscle.
Le muscle contient :
= Un sarcolemme (fine membrane élastique qui limite à l’extérieur la fibre
musculaire striée.
= Des myofibrilles, orientées parallèlement les unes aux autres dans l’axe du muscle.
Ces myofibrilles sont groupées en sarcomères, délimitées par la strie Z.
Sarcolemme
Myofibrilles
Z Z Z
Sarcomère
Fibre musculaire
= Un cytoplasme, ou sarcoplasme, contenant, outre les myofibrilles, des inclusions
de
glycogène et de lipides, la myoglobine et les organites cellulaires comme le réticulum (lieu de
synthèse des protéines) et les mitochondries.
On trouve également dans le sarcoplasme des citernes transversales séparées du système
tubulaire longitudinal, mais reliées entre elles par un système appelé triade. Ces citernes sont
destinées à contenir le Ca++ indispensable pour la contraction musculaire.
Mitochondries
Z
Tubules Citernes
transversales
2-Protéines musculaires
2-1 Myofilaments

3
Les filaments comprennent les filaments de myosine, épais, libres au milieu du sarcomère, et
les filaments d’actine, plus fins, fixés à la membrane du sarcomère (strie Z)
Actine
Myosine
Z Sarcomère Z
2.1.1 La myosine présente un corps et une tête, grossièrement semblable à un club
de golf. La tête représente environ le 1/6 ème de la longueur totale. La partie céphalique ou
méromyosine lourde est le siège de l’activité enzymatique de la myosine (capacité d’hydrolyser
l’ATP). Le segment distal, ou queue, présente une affinité pour les molécules de myosine voisines
(méromyosine légère).
Parties céphaliques
Filament de myosine
Méromyosine
légère
La jonction entre la « tête » et la « queue » est mobile, donnant ainsi à la tête la capacité de
s’orienter.
Rotation céphalique
Lors de la contraction musculaire la tête se fixe au ligament d’actine, puis opère sa
rotation (environ 45°)
2.1.2 L’actine présente la forme d’une double hélice formée de deux chaînes
monomèriques
2-2 Protéines régulatrices de la contraction musculaire
Outre l’actine et la myosine, la contraction musculaire nécessite la présence de deux autres
protéines, la troponine et la tropomyosine.

4
2.2.1 La tropomyosine est formée d’un filament fin qui s’enroule autour de l’actine
dans le sillon formé par la double hélice. Un filament de tropomyosine concerne 7 monomères
d’actine.
2.2.2 La troponine est une protéine globulaire qui chevauche la molécule de
tropomyosine vers l’une de ses extrémités. Cette protéine présente un site de fixation pour le Ca++.
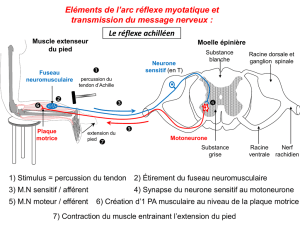
CONTRACTION MUSCULAIRE
La contraction musculaire est le résultat :
De l’activation de la plaque motrice
De la fixation du Ca++ sur la troponine
De l’activation de la tropomyosine
De la fixation de la myosine sur l’actine
De l’hydrolyse de l’ATP, responsable de la mobilité de la tête de myosine
Au repos
La plaque motrice est polarisée à - 90mV
Le calcium est enfermé dans les citernes cytoplasmiques
La troponine inhibe, via la tropomyosine, toute liaison de l’actine et de la myosine
De l’ATP est fixé sur la myosine
Les deux filaments sont libres et dans l’incapacité d’établir des liaisons.
Excitation
L’excitation du moto neurone alpha est responsable d’une activation de la plaque motrice.
L’activation se fait par libération de quanta d’acétylcholine (chaque quantum représente 0,5 mV).
Le potentiel de plaque motrice est obtenu pour des stimulations présynaptiques modérées. Il
se déclenche à - 60mV au bout de0,5 ms, et dure 10 à 20 ms. Il se propage seulement de quelques
millimètres au delà de la plaque. Si la stimulation est plus forte on observe un potentiel d’action
motrice (inversion de la polarité à + 30 mV, d’une durée plus courte (3 ms). Sa propagation est rapide
5 mètres/seconde.
Cette activation libère le Ca++ contenu dans le système cisternal du fait de l’ouverture des
canaux calciques.
Le calcium se lie à la troponine, libérant ainsi son inhibition vis à vis de la tropomyosine.
Cette activation Troponine/tropomyosine modifie le rapport Actine/tropomyosine (le ligament de
tropomyosine se déplace vers l’axe de la rainure formée par les deux brins de l’actine).
Les monomères d’actine sont alors libérés de leur inhibition, les têtes de myosine peuvent se
fixer sur l’actine.
Stimulation du
motoneurone alpha
Acétylcholine

5
Plaque motrice
Ca ++ Ca++ Mg++
Mg++ Ca++ Ca++
Troponine
Tropomyosine
Inhibition Libération du site
L’inhibition est levée
Contraction
= L’ATP qui était déjà fixé sur les molécules de myosine sous une forme dite
« énergétisée » (Mg+-ATP) provoque la brusque rotation des têtes de la myosine.
= L’hydrolyse de l’ATP fournit l’énergie nécessaire à la contraction.
= Le filament d’actine glisse le long de celui de myosine, rapprochant ainsi deux
stries Z.
= Les sarcomères sont raccourcis quasiment instantanément et les éléments élastiques
entre la sarcomère et les os sont brusquement étirés.
= Une plaque motrice concernant plusieurs sarcomères, et un neurone plusieurs
plaques motrices, la contraction peut se développer dans l’ensemble du muscle.
Plaques motrices
Z Z
Relaxation Contraction
Relaxation
L’ATP présent dans la cellule se fixe sur les têtes de myosine, séparant ainsi l’actine de la
myosine. Le calcium est repompé de façon active dans les citernes cytoplasmiques grâce à une
protéine, la calciquestrine. Ce phénomène est consommateur d’énergie, un ATP pour 2 Ca++.
La diminution de la calcicytie (calcium cytoplasmique) redonne à la troponine sa
configuration initiale, inhibitrice de la tropomyosine.
 6
7
8
9
10
11
12
13
14
15
6
7
8
9
10
11
12
13
14
15
1
/
15
100%