Digestion - Détoxification - CHUPS – Jussieu

Université Pierre et Marie Curie
Digestion - Détoxification
Niveau DCEM1
2003 - 2004
Pr. A. Raisonnier ([email protected])
Mise à jour : 21 janvier 2004

2/163 Digestion - Détoxification - Pr A. Raisonnier 2003 - 2004

Plan du cours
2003 - 2004 Digestion - Détoxification - Pr A. Raisonnier 3/163
Plan du cours
3Plan du cours
9Objectifs
11 Partie I : Aliments indispensables
13 Chapitre 1 : Définitions
14 1.1 Aliment
15 1.2 Nutriment
16 1.3 Essentiel
17 1.4 Indispensable
18 1.5 Oligoélément
19 1.6 Oligoéléments (besoins)
20 1.7 Glucides indispensables
21 1.8 Acide ascorbique = vitamine C
23 Chapitre 2 : Acides aminés indispensables
24 2.1 Acides aminés indispensables : définition
25 2.2 Acide aminé limitant : exemple des haricots verts
26 2.3 Acide aminé limitant : exemple du maïs
27 2.4 Acide aminé limitant : exemple des cacahuètes
28 2.5 Acide aminé limitant : exemple du poisson
29 2.6 Acide aminé limitant : exemple du lait de vache
31 Chapitre 3 : Acides gras indispensables
32 3.1 Lipides indispensables
33 3.2 Acide linoléique
34 3.3 Arachidonate
35 3.4 Linolénate
36 3.5 Δ-9 désaturase
37 3.6 Δ-12 désaturase
38 3.7 Δ-15 désaturase
39 3.8 Δ-6 désaturase
40 3.9 Δ-5 désaturase
41 3.10 Δ-4 désaturase
42 3.11 Famille n-9

Plan du cours
4/163 Digestion - Détoxification - Pr A. Raisonnier 2003 - 2004
43 3.12 Famille n-7
44 3.13 Famille n-6
45 3.14 Famille n-3
47 Chapitre 4 : Vitamines et coenzymes
48 4.1 Vitamines
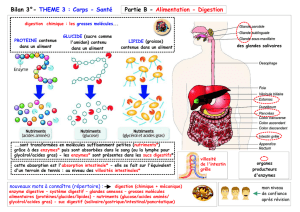
51 Partie II : Digestion
53 Chapitre 5 : Définitions
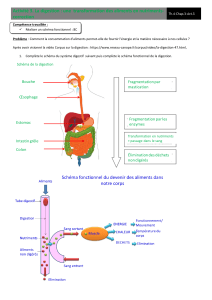
54 5.1 Digestion
55 5.2 Digestion (schéma général)
57 Chapitre 6 : La salive
58 6.1 Salive : composition
59 6.2 α-Amylase
60 6.3 Lactoperoxydase
61 6.4 Rhodanèse
63 Chapitre 7 : Le suc gastrique
64 7.1 Suc gastrique : composition
65 7.2 Mucine
66 7.3 Gradient d’acide
67 7.4 Pepsine
68 7.5 ATPase H+/K+
69 7.6 Chymosine (rennine)
71 Chapitre 8 : La bile
72 8.1 Bile hépatique : composition
73 8.2 Acide cholique
74 8.3 Acide glycocholique
75 8.4 Acide taurocholique
76 8.5 Choloyl-CoA Gly transférase
77 8.6 Acide chénodésoxycholique
78 8.7 Acide glycochénodésoxycholique
79 8.8 Acide taurochénodésoxycholique
80 8.9 Acide désoxycholique
81 8.10 Acide glycodésoxycholique

Plan du cours
2003 - 2004 Digestion - Détoxification - Pr A. Raisonnier 5/163
82 8.11 Acide taurodésoxycholique
83 8.12 Acide lithocholique
84 8.13 Acide ursodésoxycholique
85 8.14 Acide sulfolithocholique
86 8.15 Sulfotransférase (sels biliaires)
87 8.16 Triangle de SMALL et DERVICHIAN
88 8.17 Structure spatiale d’un acide biliaire
89 8.18 Micelles
90 8.19 Cycle entérohépatique des sels biliaires
93 Chapitre 9 : Le suc pancréatique
94 9.1 Suc pancréatique : composition
95 9.2 α-Amylase
96 9.3 Lipase pancréatique
97 9.4 Absorption des lipides
98 9.5 Monoglycéride acyltransférase
99 9.6 Interface lipase:colipase
100 9.7 Phospholipases
101 9.8 Cholestérol estérase
102 9.9 Trypsine
103 9.10 Activation du trypsinogène
104 9.11 α-Chymotrypsine
105 9.12 Activation du chymotrypsinogène
106 9.13 Carboxypeptidase A
107 9.14 Carboxypeptidase B
108 9.15 Peptidase A
109 9.16 Peptidase E
110 9.17 Ribonucléase A
111 9.18 Désoxyribonucléase I
113 Chapitre 10 : Enzymes intestinales
114 10.1 Maltase
115 10.2 Lactase
116 10.3 Saccharase
117 10.4 Leucine aminopeptidase
118 10.5 Prolinase
119 10.6 Prolidase
120 10.7 Phosphatase alcaline
121 10.8 Récepteurs d’absorption
122 10.9 Selles
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
1
/
163
100%