1S Devoir commun 2017 CORRECTION PARTIE I

1S Devoir commun 2017 CORRECTION
PARTIE I- RESTITUTION ORGANISEE DES CONNAISSANCES : / 10 points
Les mutations sont la source aléatoire de la diversité des allèles, fondement de la biodiversité.
Toutefois, elles peuvent aussi être à l'origine de graves maladies.
(1)Après avoir présenté l'origine des mutations, (2) discutez des conséquences plus ou moins graves de
celles-ci.
Votre exposé sera constitué d’un texte clair et structuré.
Introduction :
(Enoncé de la problématique, annonce du plan, définition des mots clés.)
Les mutations sont des phénomènes relativement rares, accidentels et aléatoires qui modifient le génome
des individus en modifiant la séquence de nucléotides de l’ADN.
Ces modifications sont à l’origine des différentes versions des gènes : les allèles qui permettent une
diversification des caractères et donc du vivant. Cependant elles sont aussi à l’origine de
dysfonctionnement et de maladies.
En fonction des cellules, des gènes touchés ou de l’étendue des modifications provoquées, les conséquences
de ces mutations vont être plus ou moins graves.
On s’interroge sur l’origine des mutations et la gravité de leurs conséquences.
Pour cela nous allons dans un premier temps expliquer l’origine des mutations, puis étudier les différentes
conséquences possibles.
Développement : (le plan est suggéré dans l’énoncé)
I – Origine des mutations :
Au cours du cycle cellulaire, pendant l’interphase, l’ADN est fidèlement recopié pendant la phase S
pour assurer le maintient du patrimoine génétique des cellules d’une génération cellulaire à la suivante.
Cette réplication est assurée par une enzyme l’ADN polymérase qui recopie chaque brin de l’ADN en
plaçant face à chaque nucléotide le nucléotide complémentaire (A-T ; C-G)
Pendant cette réplication, L’ADN polymérase peut commettre des erreurs spontanées et rares :
On peut distinguer plusieurs types de mutations, touchant un ou plusieurs nucléotides :
La fréquence des mutations est augmentée par l’action d’agents mutagènes : UV, tabac, alcool,
amiante, radiations… ces agents de l’environnement peuvent endommager l’ADN et provoquer des
modification de la séquence de nucléotides.
NB : L’ADN peut aussi être endommagé en dehors de la réplication, par exemple par les virus, qui au cours de leur
cycle de développement vont intégrer leurs gènes au génome de la cellule infectée et modifier celui-ci.
Le plus souvent l’erreur est détectée et réparée par des systèmes enzymatiques de contrôle qui
vérifient les différentes étapes du cycle cellulaire (endonucléases) qui éliminent la portion contenant la
mauvaise séquence, l’ADN polymérase re-synthétisant la partie manquante.
Si ces erreurs ne sont pas corrigées et si les modifications n’empêchent pas la survie de la cellule, il
apparaît donc une mutation.

II – Conséquences plus ou moins graves des mutations :
1. Des conséquences différentes en fonction de la modification de la protéine codée.
Un gène est une séquence de nucléotides qui contient les informations nécessaires à la synthèse d’une
protéine. La séquence de nucléotides code pour une séquence d’acides aminés en fonction d’un code quasi
universel : le code génétique qui fait correspondre un codon (séquence de 3 nucléotides) à un acide aminé.
Si la séquence du gène change, cela peut modifier la séquence d’acides aminés de la protéine qui peut alors
voir sa forme modifiée, ce qui modifie sa fonction.
Ainsi les substitutions peuvent se traduire par des séquences d’acides aminés plus ou moins
différentes ce qui va modifier la fonction de la protéines sauf si le nouveau codon apparu code pour le
même acide aminé grâce à la redondance du code génétique.
Les insertions et additions entrainent un décalage du cadre de lecture qui est susceptible de modifier plus
profondément les séquences ou de faire apparaître un codon stop qui interrompt de façon anticipée la
synthèse de la protéine : écourtée, elle aura peu de chance d’être fonctionnelle.
NB : la région de la protéine qui sera touchée est importante une modification même ponctuelle de la protéine, peut
avoir de graves conséquences si c’est la zone active de la protéine qui est modifiée.
Cette modification du phénotype moléculaire peut modifier le phénotype cellulaire qui à son tour peut
modifier le phénotype macroscopique = le dysfonctionnement des organes provoqué sera responsable des
symptômes d’une maladie.
2. Des conséquences différentes selon les cellules touchées :
Une mutation peut survenir dans une cellule somatique (cellules du corps ≠ des cellules
reproductrices) et est alors présente dans toutes les cellules filles issues de la division de cette cellule si elle
ne provoque pas la mort de la cellule. Cette mutation pourra avoir des conséquences sur le fonctionnement
de la cellule et modifier plus ou moins gravement son activité mais elle disparaîtra avec l’individu à sa
mort.
Si les protéines touchées sont les protéines qui régulent le cycle cellulaire, la cellule peut devenir
cancéreuse : elles se multiplie de façon anarchique, perd sa différenciation, constituant une tumeur et peut
acquérir une mobilité se traduisant par la formation de métastases.
Cancérisation++
Mais elle peut intervenir aussi dans une cellule germinale (cellules reproductrice) elle sera alors
transmise à la descendance et devient héréditaire.
Ces mutations sont responsables de l’apparition de nouvelles versions de gènes : les allèles qui pourront
faire apparaître des phénotypes alternatifs avantageux, sélectionnés par l’évolution et ces nouveaux allèles
se répandront dans les populations. La combinaison des mutations (le hasard) et de la sélection de
l’environnement (la nécessité) est responsable de l’évolution des populations et des espèces.
Par exemple dans les populations de bactéries, des mutations peuvent faire apparaître des résistances aux
antibiotiques.
Erreur%de%l’ADN%polymérase%
Agents%mutagènes%
%

Mais ces mutations peuvent aussi être responsables des maladies génétiques, héréditaires.
La transmission des allèles mutés au cours de la reproduction sexuée transmet le codage d’une protéine
anormale, responsable de dysfonctionnements.
Par exemple la mucoviscidose est une maladie génétique assez répandue, elle est due à une mutation du
gène CFTR qui code pour une protéine (CFTR) assurant la régulation de la fluidité du mucus.
L’allèle muté code pour une protéine modifiée qui n’assure plus cette régulation, les cellules sécrétrices
produisent un mucus trop épais qui s’accumule, favorise les infections et obstruent différents canaux
(bronches, intestins, canaux de production des enzymes digestives, canaux reproducteurs) entraînant de
graves problèmes respiratoires, digestifs, reproducteurs…
Mutation+somatique+ou+germinale+
Dysfonctionnement+des+
cellules+épithéliales+:++

Conclusion
Les mutations sont donc le résultat d’erreurs de l’ADN polymérase au cours de la réplication, de l’action
d’agents mutagènes, non corrigée par les systèmes de surveillance cellulaire.
Elles sont source de diversification du vivant, elles permettent l’apparition de nouveaux allèles qui codent
pour des protéines alternatives responsables de phénotypes alternatifs qui seront soumis au filtre sélectif
de l’environnement.
Elles peuvent être responsables de maladies graves : cancers si les cellules somatiques sont atteintes,
héréditaires si les cellules germinales sont touchées.
Ouvertures possibles:
- L’impact d’une modification du génome sur la santé n’est pas un déterminisme absolu. Il existe des mutations qui
rendent plus probable le développement d’une maladie sans pour autant la rendre certaine. En général les modes de vie
et le milieu interviennent également, et le développement d’une maladie dépend alors de l’interaction complexe entre
facteurs du milieu et génome. (Ex : diabète de type II)
- Comment lutter contre les conséquences des maladies génétiques : prévision, dépistage, thérapie génique ?
PARTIE II/1- RAISONNEMENT SCIENTIFIQUE A L’AIDE DES CONNAISSANCES / 5 POINTS
1. : B/C
2. : A/D
3. : A/C/D
4. : B
5. : B/D
PARTIE II/2 RESOUDRE UN PROBLEME / 5 POINTS
A partir de l’analyse des documents fournis, proposez un scénario pour l’histoire des gènes
codant pour les pigments visuels. Vous compléterez le document 4 en plaçant l’apparition des
différents gènes.
PB posé
Infos :
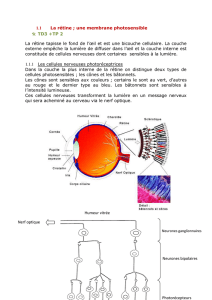
Document 1- Les gènes des opsines chez l’Homme.
Chez l’Homme, les opsines (= protéines
= pigments visuels) contenues dans les
trois types de cônes (= cellules de la
rétine, photoreceptrices, contiennent
chacune un type de pigment) sont les
produits de trois gènes localisés sur deux
chromosomes différents.
Infos pertinentes
Connaissances
Déductions
3 gènes ≠, (locus ≠)
sur 2 chromosomes
≠ codent pour 3
opsines (pigments
rétiniens) ≠ : M, L, S
Opsines = protéines contenues
dans les cônes de la rétine :
cellules photosensibles
spécialisées dans la perception
des couleurs.
Il existe 3 types de cônes
contenant chacun 1 opsine
Chez l’homme il existe 3 types de
cellules photoreceptrices percevant les
couleurs : les cônes qui contiennent
chacune un type de pigment visuel
(opsine) M ; L ou S
Ces opsines sont codées par 3 gènes ≠
situés sur 2 chromosomes ≠.
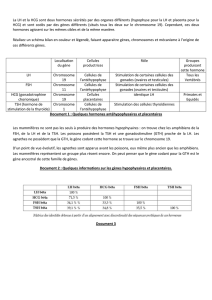
Document 2- Tableau des identités (exprimées en %) obtenu à partir d’une comparaison des
séquences nucléotidiques des gènes des opsines S, M et L .
Gène opsine M
Gène opsine L
Gène opsine S
Gène opsine M
100
Gène opsine L
96 (MAX)
100
Gène opsine S
44(MIN)
43 (MIN)
100

Infos
pertinentes
Connaissances
Déductions
Fort taux
d’identité (>
40%)
Max
d’identités :
entre M et L :
96%
Min entre S et
(M, L) : ± 45%
La comparaison des identités
permet d’établir le lien de
parenté entre les gènes
étudiés
Plus le % d’identité est élevé
plus le lien de parenté est
étroit, plus les gènes
comparés partagent un gène
ancestral commun récent.
Ce gène ancestral a subi une
duplication et une
transposition suivie de
mutations indépendantes. =>
famille multigénique
Le fort % d’identités entre les 3 gènes témoigne
d’un lien de parenté.
- M et L partage le plus grand% d’=, ils ont le
lien de parenté le plus étroit : ils proviennent
du gène ancestral le plus récent
- (M,L) et S partage un gène ancestral plus
ancien.
- M, L et S proviennent de 2 duplications
successives :
1er ! S et le gène ancestral de M et L avec ne
translocation sur un autre chromosome (7!X)
2ième ! M et L (tous 2 sur le chromosome X)
ces 3 gènes constituent une famille
multigénique
Document 3 - Gènes des opsines et vision chez les primates.
Infos pertinentes
Connaissances
Déductions
Chez les primates de
l’ancien monde, 3 gènes !
trichromatie (vision de 3
couleurs : R, V, B) sur 2
chromosomes ≠ (7, X)
Chez les primates du
nouveau monde : 2 gènes
! dichromatie (vision de 2
couleurs (B, V) sur 2
chromosomes ≠ (7, X)
Les différentes opsines
permettent la vision de
couleurs différentes
Toutes les espèces ne perçoivent pas les
couleurs de la même façon, cela dépend des
gènes des opsines qu’ils possèdent.
Chez les primates, on distingue ceux de
l’ancien monde qui sont trichromates grâce
à la possession des 3 gènes
Et ceux du nouveau monde qui sont
dichromates car ils ne possèdent que 2 gènes
On peut faire l’hypothèse que la 2ième duplication donnant les gènes M et L ne s’est pas déroulée chez les
primates du nouveau monde.
L'Homme et certains autres Primates sont trichromates c'est-à-dire que leur vision des couleurs est
due à la présence de trois catégories de pigments contenus dans les cônes.
Les singes de l'Ancien Monde (Bonobo, Chimpanzé, Gorille et Macaque) présentent le gène S
(codant l’opsine bleue) sur le chromosome 7 et les gènes L et M (codant respectivement les opsines
rouge et verte) sur le chromosome X.
Mais la plupart des mammifères sont le plus souvent dichromates: leur vision dépend de deux
catégories de pigments. Ainsi les singes du Nouveau Monde (Amérique) possèdent le gène S sur
un autosome quelconque et un seul autre gène nommé M/L codant pour une autre opsine sur le
chromosome X
Gène%de%
l’opsine
%M/L%
 6
7
6
7
1
/
7
100%