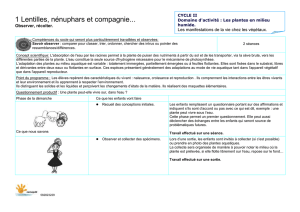
Transfert d`un insecticide systémique, l`imidaclopride, chez la tomate

N° d’ordre : 2543
THESE
présentée
pour obtenir
LE TITRE DE DOCTEUR DE L’INSTITUT NATIONAL POLYTECHNIQUE DE TOULOUSE
École doctorale : SEVAB
Spécialité : Qualité et Sécurité des Aliments
Par
Haifaa AL–SAYEDA
Transfert d’un insecticide systémique,
l’imidaclopride, chez la tomate : implication du
transport phloémien
Soutenue le 15 novembre 2007 devant le jury composé de :
Thierry LAMAZE Président
François LAURENT Directeur de thèse
Maria Louisa FERNANDEZ CRUZ Rapporteur
Patrick RAVANEL Rapporteur
François LAURENT Examinateur

REMERCIEMENTS
Je tiens à exprimer ma reconnaissance à Monsieur Jacques Tulliez et à Monsieur Jean-Pierre
Cravedi pour m’avoir accueillie dans l’UMR des Xénobiotiques ainsi que pour leur soutien tout
au long de ce travail.
Je remercie Madame Maria Louisa Fernandez Cruz et Monsieur Patrick Ravanel d’avoir accepté
d’être rapporteurs de mon travail de thèse et Monsieur Thierry Lamaze et Monsieur François
Laurent d’avoir accepté d’être examinateurs de mon travail de thèse.
Je tiens à remercier Monsieur François Laurent, mon directeur de thèse de m’avoir donné
l’opportunité de travailler sur un tel sujet et pour sa confiance en moi au travail, ses précieux
conseils et son soutien.
Je remercie Madame Sophie Lorber pour son soutien et son aide, particulièrement lors de la
rédaction.
Je remercie Madame Estelle Rathahao et Monsieur Laurent Debrauwer pour la réalisation des
différentes analyses en spectrométrie de masse et leurs explications avisées.
Un grand merci à l’ensemble de mes collègues de l’UMR des Xénobiotiques, en particulier
Roselyne, Sabrina, Bérengère et Chary, pour l’aide et la sympathie qu’ils m’ont témoignées
durant ces quelques années passées ensemble.
Je remercie vivement le Chef du Département de la protection des plantes de la Faculté du Génie
Agricole de l’Université de Damas et le Ministre des Etudes Supérieures pour leur confiance et
pour le financement de ma thèse.
Je remercie Monsieur Mohamad Jamal al Hajjar de m'avoir permis d'effectuer mes études en
France.
Un grand merci à tous les professeurs du Département de la Protection des Plantes, auxquels je
dois ma formation. Je tiens à remercier tout particulièrement Madame Houda Kawas pour son
soutien. Je remercie vivement l'ensemble du personnel de ce Département pour leur sympathie.
Je voudrais exprimer toute ma gratitude à ma mère et à mon père pour la confiance qu’ils me
témoignent depuis toujours et pour leurs encouragements.
Je remercie infiniment mes sœurs Bashirah, Wafaa, Rajaa, et mes frères Nemer, Mohammad,
Ahmad, Mahmoud et Maher et aussi leurs enfants pour leur grand amour.
Je tiens à remercier tout particulièrement mon fiancé Saleh, pour son amour et sa patience.
Enfin, je remercie mes amies de Syrie, Safaa, Nadya, Bouthena et Mervat pour leur soutien ainsi
que tous les amis que j’ai connues à Toulouse et à Nancy pour leur sympathie et leurs
encouragements.

PUBLICATION DANS UNE REVUE INTERNATIONALE A COMITE DE LECTURE
Haifaa AL-SAYEDA, Sophie PASCAL-LORBER, Chary NALLANTHIGAL, Laurent
DEBRAUWER, François LAURENT. Fate of [14C] imidacloprid in tomato plants after soil
treatment. Environ Chem Lett, DOI 10.1007/s10311-007-0121-2.
POSTERS PRESENTES DANS DES COLLOQUES
Haifaa ALSAYEDA, Sophie PASCAL-LORBER, Jean-Philippe BESSE, François
LAURENT (poster). Uptake by edible plants of 4-n-Nonyl-phenol and Diuron from various
types of sludge used as soil improvement. SETAC, The Hague (Netherland), 7-11 May 2006.
Haifaa ALSAYEDA, Sophie PASCAL-LORBER, François LAURENT (communication
orale). Transfert des résidus de pesticides vers les fruits : étude d’un modèle, l’imidaclopride,
chez la tomate. XXXVI GFP, Strasbourg, 29-31 mai 2006.

ABREVIATIONS
2,4-D : acide 2,4-dichlorophénoxyacétique
6–CAN : acide 6-chloronicotinique
ARfD : acute reference dose, dose aiguë de référence
ARTAC : Association pour la Recherche Thérapeutique Anticancéreuse
CL50 : concentration létale 50, utilisée pour la concentration d'une
substance toxique létale pour la moitié des animaux exposés
Cyt P-450 : enzyme à cytochrome P-450
DDT : dichlorodiphényltrichloroéthane
DG SANCO : Direction Générale « santé et protection des consommateurs »
DGCCRF : Direction Générale de la Concurrence de la Consommation et de
la répression des fraudes
DJA : dose journalière admissible
EPA : Environmental Protection Agency
FAD : flavine adénine dinucléotide
FMN : flavine mononucléotide
H' : constante de la loi de Henry
INSERM : Institut National de la Santé et de la Recherche Médicale
JMPR : joint meeting on pesticide residues
Kd : adsorption d’un composé sur la matière organique
Kom : adsorption sur la matière organique
Kow : coefficient de partage octanol/eau
LD50 : dose létale 50, désigne la dose entraînant la mort de 50% de la
population examinée
LMR : limite maximale de résidus
M01 (WAK 4103) : composé 5-hydroxy
M06 (WAK 3745) : oléfine
M09 (WAK 4140) : composé dénitré
MSA : Mutualité Sociale Agricole
NAD : nicotinamide adénine dinucléotide
NADPH : nicotinamide adénine dinucléotide phosphate
NOEL : no observed effect level, ou dose sans effet

OCP : pesticides organochlorés
OMS : Organisation Mondiale de la Santé
Pa : pascal, unité internationale de mesure de pression
pKa : pH auquel 50% des groupes d’un acide ou d’une base sont
ionisés
POP : produit organique persistant
RCF : facteur de concentration dans les racines
TCPY : 3,5,6-trichloro-2-pyridinol
Tm : point de fusion en degré Celsius
TRR : taux de radioactivité résiduel
TSCF : facteur de concentration dans le flux de transpiration
VP : pression de vapeur
WS : solubilité dans l’eau
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
1
/
174
100%