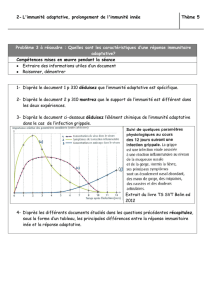
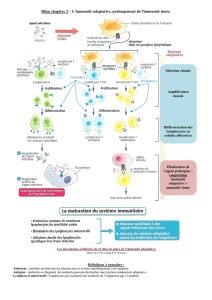
UE9 - Immunologie Mécanismes effecteurs de l`Immunité Innée

UE9 - Immunologie
Date : 11/03/2016 Horaire : 10h45/12h45
Promo : PCEM2 2015/2016 Enseignant : Jean-Jacques HOARAU
Ronéistes :
VILLARET Chloé dit « la princesse »
GUBEREK Arnaud
Mécanismes effecteurs de l’Immunité Innée
I. Introduction.
A. Notions de base.
B. Cellules, organes et tissus de l’immunité.
II. Les pathogènes.
III. Deux systèmes immunitaires (inné, acquis).
IV Les mécanismes anti-pathognes de l’immunité innée.
A. Les barrières épithéliales.
B. Les substances chimiques non spécifiques (peptides antimicrobiens)
C. Les cellules effectrices non spécifiques (fin de ce ronéo mais le cours
continuera sur cette sous partie).

I. Introduction.
A. Quelques notions de bases.
- Immunologie: L’étude du système immunitaire et de ses réponses contre les micro-organismes invasifs.
On a l’impression que cette description limite le rôle du système immunitaire à la lutte contre les
infections, mais il assure aussi d’autres fonctions (au niveau des cellules tumorales, sur les cellules,
molécules dangereuses pour l'organisme)
- Immunité: du latin « Immunis », signifie « dispensé de, exempté de » ; indique un état de protection
contre telle ou telle maladie infectieuse. La plus ancienne référence remonte à̀ -430 avant JC, épidémie
de peste à Athènes.
- Système Immunitaire (SI): Ensemble des cellules, tissus, et des molécules qui concourent à̀ opposer
une résistance aux infections. L'atteinte du SI peut entraîner des immunodéficiences, des maladies auto-
immunes etc.
Rappels : bien que l’on décrive deux facettes de l’immunité (acquise-spécifique et innée), il ne faut pas se
dire que ces deux mécanismes fonctionnent séparément : ils sont complémentaires, et il n’y a qu’une
immunité.
Fonctions physiologiques du système immunitaire :
Toutes ces fonctions sont assurées autant par le SI inné que le SI adaptatif. Le système immunitaire est
capable de reconnaître tout ce qui est étranger à l’organisme, pour maintenir l’intégrité de ce dernier. Donc
la notion importante à retenir est que le SI n'a pas une action qu'au niveau des infections mais aussi au
niveau des greffons par exemple, ou même des cellules tumorales.
B. Cellules, organes et tissus de l’immunité.
Rôle du système immunitaire
Implications
Défense contre les infections.
Un déficit immunitaire entraîne une aggravation
de la sensibilité aux infections ; par exemple dans
le SIDA.
La vaccination stimule les défenses immunitaires
et protège contre les infections.
Le système immunitaire reconnaît les
greffons tissulaires et les protéines
nouvellement introduites et y répond.
Les réponses immunitaires sont des barrières
importantes à la transplantation et à la thérapie
génique.
Défense contre les tumeurs.
Possibilité d’une immunothérapie du cancer,
puisque les facteurs présentés sur les cellules
tumorales sont reconnus comme « étrangers ».

Les cellules immunocompétantes sont à la base de l’immunité, et dérivent toutes de CSH (Cellules Souches
Hématopoïétiques) se trouvant dans la moelle osseuse (à partir du 7ème mois de vie utérine, puisqu’on en
trouve dans le sac vitellin, chez l’embryon, dans le foie et la rate chez foetus).
Les CSH représentent seulement une cellule sur 50 000 dans la moelle osseuse (stable au long de la vie, sauf
en cas d’infection où ce nombre peut augmenter de façon transitoire). Particularité de ces CSH, elles sont
capables d'auto-renouvellement, de se diviser mais leur nombre reste stable sauf dans certaines situations
où on a besoin d'un nombre plus grand en cellule immunitaire ( lors d'infection par exemple).
Les deux lignées de descendance des CSH sont la
lignée lymphoïde (immunité adaptative principalement LyT lyB, sauf NK dérivant de cette lignée
mais qui font parti de l'immunité inné).
la lignée myéloïde qui est à l'origine de la majeure partie des cellules du SI inné.
Attention même si les lymphocytes NK dérivent de la lignée lymphocytaire ils font partie de l’immunité
innée et non adaptative. Donc lignée lymphocytaire ne veut pas dire immunité adaptative (car il y a les
lymphocytes NK).
Au départ les progéniteurs (cellules souches myéloïdes et lymphoïdes) sont des cellules circulantes mais ils
gagnent rapidement les organes lymphoïdes secondaires ou les tissus périphériques pour devenir des
cellules résidentes et il ne reste que très peu de progéniteurs circulants. Ils n’ont pas de capacité de
renouvellement contrairement aux cellules souches et on compte environ 1 cellule souche myéloïde pour
10.000 cellules dans la moelle osseuse.
Les cellules souches sont multipotentes (capacité auto-renouvèlement quasi-infinie), mais cette multipotence
sera perdue avec leurs différenciations. Les différenciations se font à l’aide de facteurs de croissance
(Colony Stimulating Factors, interleukines).
Les facteurs de croissance et cytokines nécessaires à la différenciation des CSH et des progéniteurs sont
produits par les cellules stromales de la moelle osseuse (adipocytes, cellules endothéliales, fibroblastes,
macrophages, lymphocytes B).

Les plaquettes et les hématies dérivent aussi des CSH, et ne sont pas considérées comme
immunocompétentes (ce ne sont pas des cellules de l’immunité) bien que de récentes études tendent à
prouver le contraire.
Lignée lymphoïde :
NK, Th, Tc, B
Lignée myéloïde :
mono, dendritique, neutrophile, éosinophile, basophile, plaquettes, érythrocytes.
Chaque cellule a une fonction plus ou moins marquée dans un type d’immunité :
A propos des organes lymphoïdes :
Organes lymphoïdes primaires ou centraux (maturation et fabrication des cellules
immunocompétentes):
Immunité non spécifique
(Action immédiate)
Immunité spécifique
(Action retardée, mémoire)
- Monocytes
- Macrophages
- Polynucléaires
- Lymphocytes NK
- Cellules présentatrices d’antigène
CPA :
o Macrophage
o Cellule de Langerhans
o Cellule dendritiques
o Lymphocyte B
- Lymphocytes T
- Lymphocytes B

- Thymus (rôle très important dans l'éducation des LyT) malheureusement c'est un tissu qui se réduit
en taille avec l'âge, il participe à un mécanisme appelé l'immunoscénesence. Ainsi au fur et à mesure
on éduque de moins en moins bien les lyT...)
- Moelle osseuse
Organes et tissus lymphoïdes secondaires ou périphériques (là où se fait la présentation des
antigènes aux cellules lymphocytaires):
- Ganglions lymphatiques (La résidence principale des lymphocytes)
- Rate (surtout pour les LyB)
- Tissus lymphoïdes associés aux bronches
- Anneau de Waldeyer (amygdales, végétations)
- Ganglions mésentériques
- Plaques de Peyer (système digestif)
- Tissu lymphoïde urogénital
- Appendice vermiforme
Tous ces organes servent à distribuer les cellules du SI, et les rapprocher des différentes régions pouvant
être infectées.
Les cellules de l’immunité innée n’ont pas vocation à résider dans les organes lymphoïdes secondaires,
contrairement aux cellules de l’immunité adaptative.
Organes lymphoïdes secondaires sont retrouvés près des épithéliums en contact avec l’extérieur: bronches,
amygdales etc…
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
1
/
24
100%