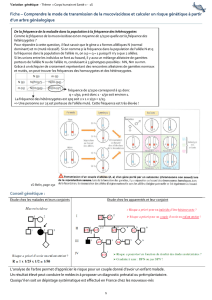
facteurs affectant l`equilibre


Étude de la diversité génétique intraspecifique
Typer 25-30 individus non apparentés
Marqueurs moléculaires

Représentation des génotypes dans un tableau (Exel)

Équilibre de Hardy-Weinberg

Mis en évidence en 1908 par G.h. Hardy et W.
Weinberg
Les fréquences alléliques et genotypiques restent
stable de génération en génération dans une
population diploïde idéale
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
6
7
8
9
10
11
12
13
14
15
16
17
18
19
1
/
19
100%