le cerveau : quel type de machine - Laboratoire Matière et Systèmes

LE CERVEAU :
QUEL TYPE DE MACHINE ?
Jean-Pierre HENRY
!"#$%&$'#()$(#$%*$#%*$(+,+#"&$(-'(./01(
2-34#-&4"#$(5-67#$($&(189&7,$9(.4,:;$<$9((
=50(>?@>(./01(A(=B"C$#9"&+(D-#"9(>(!")$#4&(
54)+#-&#"%$(E((
Rodica RAVIER
Mardi 25 novembre 2014
1-;;$(F';$9(G4#)$&H(IB96&'&(D-9&$'#(

! !"#!$#%&#'(!)#!*+,-!*+)$!.'/!0-%#!'(-%#!$1+/#2!3!
)+-%#!'&,/2!4(5()#!#/.6$#!*#!3'#$-'(&+;+:*4B"J'$(
%$B&#-;!7!/+)!%89#!#/-!*#!:!*+))#%!9'!$+;;(),$'<+)!=2!
+(!*#!9'!>',%#!'?#)*%#@!I;(BK-L4'&$(#"$B(M(%$(J'K";(#$N4"&(A!
;',/!$+;;#!-+(/!9#/!+%B')#/!.#%$#.<>/!C!#)&+,#)-!
9#(%/!*#%),#%/!.%+9+)B#;#)-/2!#-!4(#!-+(/!9#/!
;D$'),/;#/!;+-#(%/!*#!9'!;+#99#!#-!!*(!E(9E#!C!+)-!
9#(%/!%#.%D/#)-')-/!'F-%D/2!,9!$+)/<-(#!E,#)!%D#99#;#)-!
()!$#)-%#2!+G!95#H$,-'<+)!.D%,.1D%,4(#!/#!;#-!#)!
%'..+%-!'&#$!-#9!+(!-#9!;D$'),/;#!
O$B#"(G$#P94B(
I'<6%#!#-!ID;+,%#!JKLMNO!

PQRSTUVWRPTQ!
• !"XY!QXVSTQXY!UZQY!"X!WXS[XZV!
• !"Z!WT\QPRPTQ!
• !"Z!I]ITPSX!
• !"Z!WTQYWPXQWX!
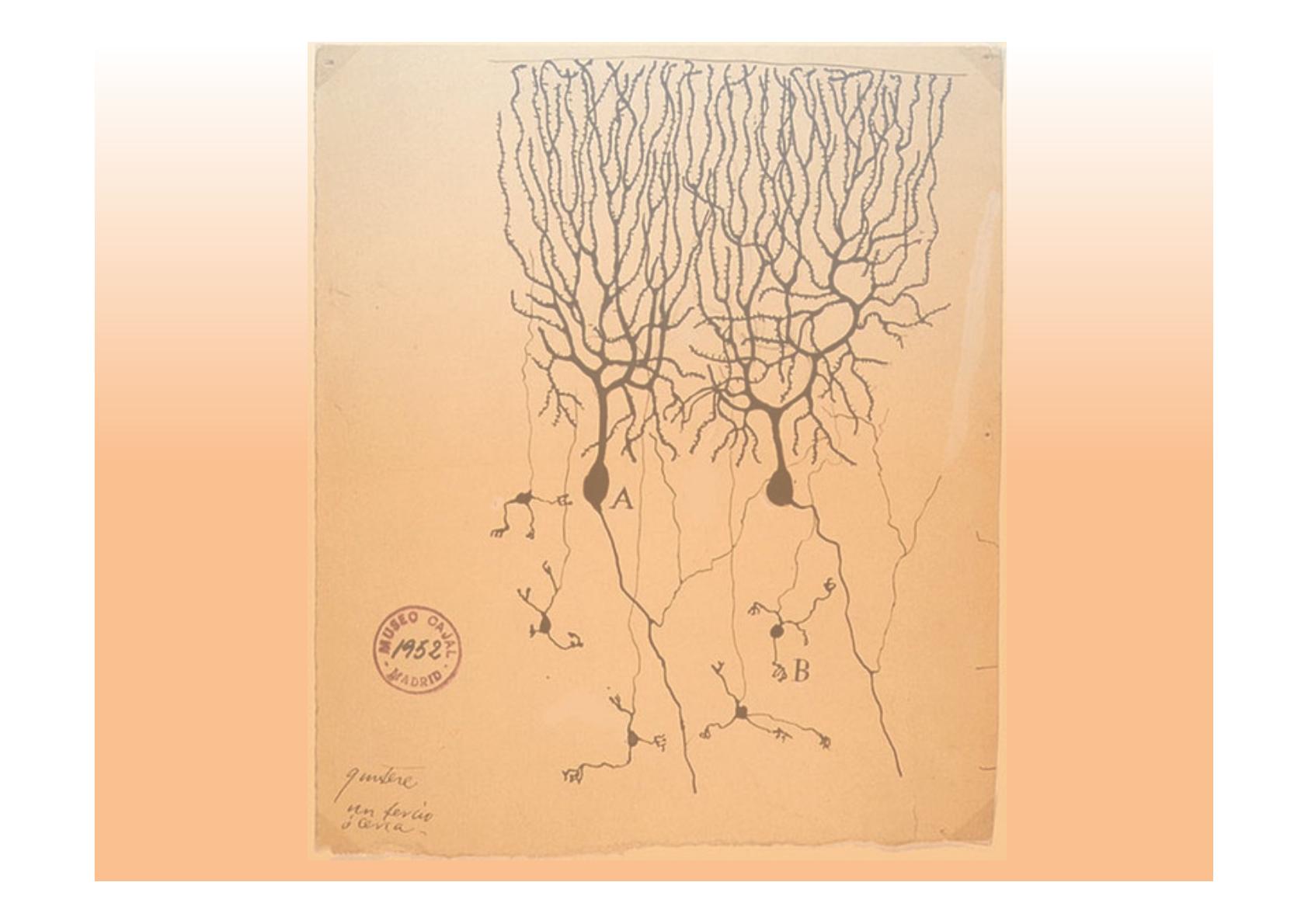
IB)'%64BE(2$9((B$'#4B$9((
U#/!$#99(9#/!.'/!$+;;#!9#/!'(-%#/!
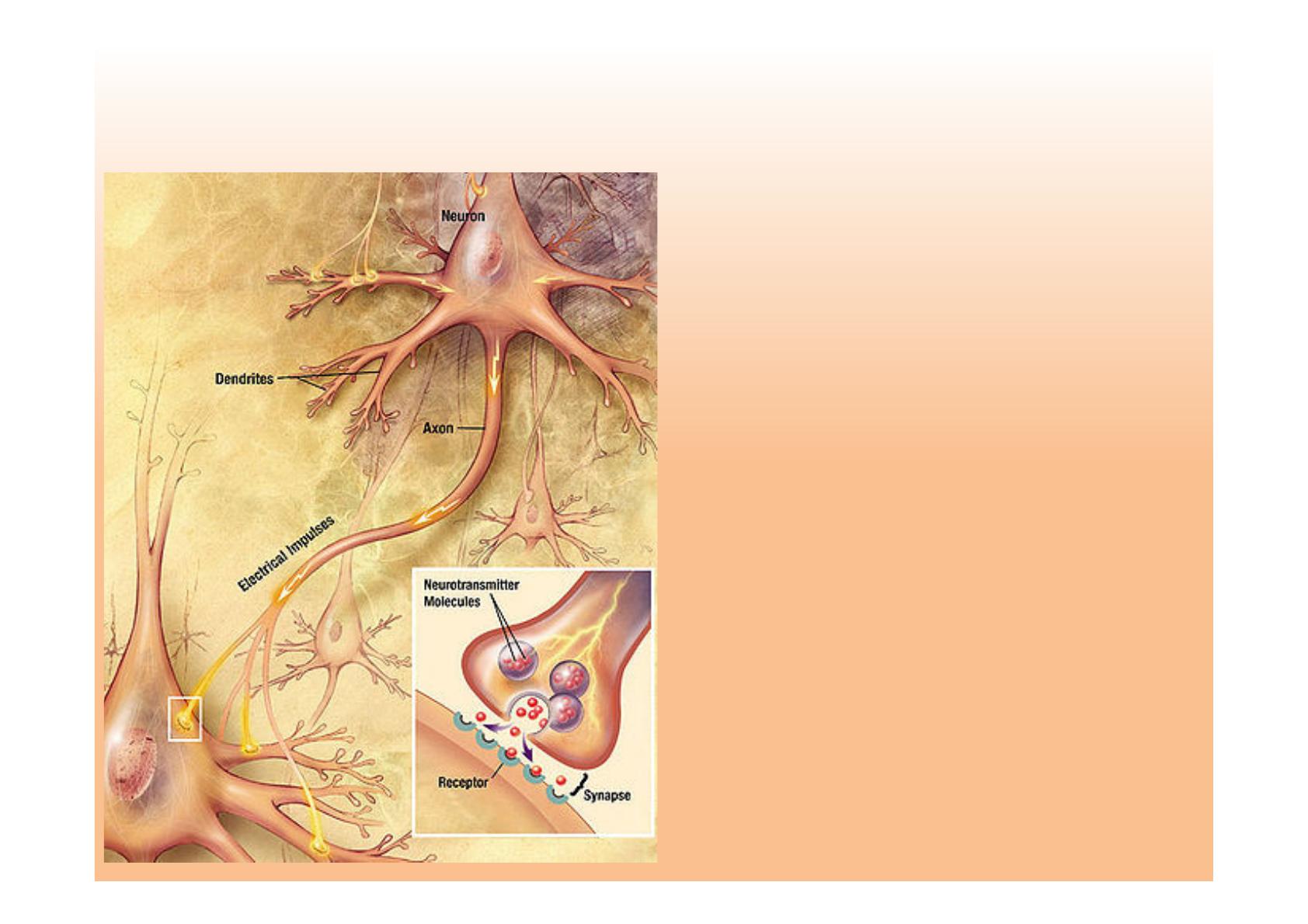
"#!)#(%+)#7!*(!;+%.1+9+B,4(#!'(!>+)$<+))#9!
• "#!)#(%+)#!.#%;#-!9'!
$,%$(9'<+)!*#!9^,)>+%;'<+)2!
'&#$!/#/!$+;.+/')-#/!
D9#$-%,4(#!#-!$1,;,4(#!
• "^#)-%D#!/#!>',-!.'%!;$9()$B)#"&$9(
'&#$!'..'%,<+)!*^()!$+(%')-!
D9#$-%,4(#!4(,!/#!.%+.'B#!9#!
9+)B!*#!;Q-<4B$!!
• Z!9^#H-%D;,-D2!9,ED%'<+)!*(!
)#(%+-%')/;#?#(%!$+)-#)(!
*')/!9#/!&D/,$(9#/!/C)'.<4(#/!
• W#9(,_$,!#)!/#!$+;E,)')-!'&#$!
*#/!#+%$:&$'#9!9+$'9,/D/!/(%!/(%!
9#/!*#)*%,-#/!J+(!9#!$+%./!
$#99(9',%#O!*D$9#)$1#!9^'..'%,<+)!!
*^()!.+-#)<#9!D9#$-%,4(#!!
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
1
/
93
100%











