Chapitre 4 L`oeil et la vision

1
Chapitre 4 L’oeil et la vision
I. L’oeil, un organe sensoriel.
Répondre aux questions p 305
1. Structure de l’œil de Mammifère.
L’oeil de Mammifère comprend :
Des structures transparentes qui vont donc laisser passer la lumière : humeur aqueuse à l’avant du
cristallin, le cristallin lui-même, humeur vitrée à l’arrière, cornée sur la face avant de l’œil.
Des systèmes optiques : le cristallin, jouant le rôle d’une lentille convergente, la pupille jouant le rôle
de diaphragme.
La rétine, recevant les images et les transformant en messages nerveux acheminés vers le cortex par le
nerf optique.
Des enveloppes protectrices : cornée à l’avant, choroïde et sclérotique dans la partie de l’œil incluse
dans le globe oculaire. Chez l’homme et de nombreuses espèces, la choroïde est recouverte d’un
épiderme riche en mélanine, substance sombre qui évite les reflets à l’intérieur de l’œil
2. Structure et fonction du cristallin.
Le cristallin est un organe transparent. Or les cellules renferment de nombreux organites, comme le
noyau et les mitochondries, obstacles à la transparence.
Les cellules de la périphérie du cristallin se divisent en repoussant les cellules plus anciennes vers le
centre. Après s’être divisées, les cellules nouvellement formées perdent leur noyau et leurs autres
organites ; leur cytoplasme se remplit de protéines particulières appelées cristallines, qui forment un
réseau transparent. Au-delà de 20 ans chez l’Homme, les cellules du cristallin ne sont plus renouvelées :
le cristallin va donc être un organe particulièrement soumis au vieillissement.
Le cristallin est rattaché à la choroïde par des ligaments suspenseurs et des muscles ciliaires qui peuvent
modifier son étirement : ainsi le cristallin a une convergence plus ou moins forte, ce qui permet de faire
en sorte qu’un objet ait son image toujours formée sur la rétine quel que soit son éloignement. Cette
modification de la convergence du cristallin s’appelle l‘accommodation.
3. Des maladies liées au cristallin.
Le cristallin, constitué de cellules hautement spécialisées et non renouvelées, supporte mal le
vieillissement : soumis en particulier à de trop fortes expositions aux ultra-violets, il s’opacifie. C’est la
cataracte, que l’on peut opérer en enlevant le cristallin et en le remplaçant par un implant.
Le vieillissement du cristallin affecte également sa souplesse. Lorsqu’il ne peut plus se déformer
suffisamment, l’accommodation devient insuffisante, empêchant la vision de près : c’est la presbytie.

2
II. Organisation de la rétine.
Répondre aux questions p 307 et 309.
1. Structure.
La rétine est constituée de plusieurs couches de cellules :
- Contre l’épiderme pigmentaire, des cellules photoréceptrices de deux types différents dans l’espèce
humaine, cônes et bâtonnets. Dans d’autres espèces animales, on ne trouve que des bâtonnets.
- Sous cette première couche, se trouve une couche de cellules bipolaires, reliant quelques cônes et/ou
bâtonnets à une cellule ganglionnaire sous-jacente.
- La dernière couche, la plus externe, est constituée de cellules ganglionnaires, neurones dont les
prolongements forment le nerf optique.
Schéma p 306.
2. Cônes et bâtonnets.
Les cônes et les bâtonnets sont des cellules photoréceptrices, à l’organisation cellulaire particulière : ils
sont constitués :
- D’un segment externe, avec de nombreux replis membranaires où se trouvent les pigments
photorécepteurs.
- D’un segment interne comprenant les organites cellulaires autres que le noyau
- Du noyau, situé à la base de la cellule.
Cônes et bâtonnets diffèrent par la forme de leur segment externe, par leur pigment photorécepteur, et
par les propriétés de ce pigment :
Cônes
Bâtonnets
Forme du segment externe
Large et court
Long et fin
Nature du pigment
photorécepteur
Opsine R, V ou B
rhodopsine
Propriétés du pigment
P 308.
Peu sensible à l’intensité
lumineuse ; sensibilité à
différentes longueurs d’onde
donc aux couleurs
Fonctionnent même pour une
faible intensité lumineuse ; pas
de vision des couleurs.
Doc. 3 p 309. Les cônes sont donc adaptés à la vision diurne. L’opsine B (bleu) capte les faibles
longueurs d’onde, l’opsine V (vert) les longueurs d’onde moyennes et l’opsine R (rouge) les grandes
longueurs d’onde.
Opsine ou rhodopsine sont constituées de l’association de deux composants : une protéine, dont la
conformation détermine la longueur d’onde captée, et une molécule de rétinal, dérivé de la vitamine A.
La rétine est un tissu nerveux, constituée d’un ensemble de cellules nerveuses interconnectées. Elle
possède la propriété de réagir par une variation de potentiel électrique à une stimulation extérieure
(lumière).
3. Répartition des photorécepteurs rétiniens
La rétine n’est pas uniforme. La fovéa, point de la rétine situé directement à l’arrière de la pupille, sur
l’axe optique du cristallin, ne contient que des cônes : elle est donc particulièrement adaptée à la vision

3
diurne. Les bâtonnets se trouvent donc sur les côtés : la vision crépusculaire est plus facile si on regarde
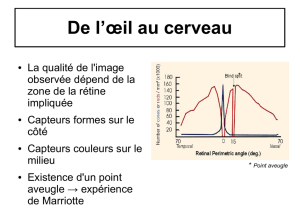
de biais. Le point aveugle ne renferme aucune cellule photoréceptrice : il correspond au point de départ
du nerf optique.
4. Une anomalie génétique : le daltonisme.
Certaines mutations ponctuelles peuvent rendre la protéine non fonctionnelle. Dans le cas des opsines, si
l’une des opsines est non fonctionnelle, la vision, normalement trichromatique, devient dichromatique :
c’est le daltonisme.
Comme deux des gènes des opsines se trouvent sur le chromosome X, le daltonisme est beaucoup plus
fréquent (8% de la population) chez les hommes que chez les femmes (0,5%).
III. Photorécepteurs et évolution.
Répondre aux questions p 311
Dans l’espèce humaine, les gènes codant pour la partie protéique des opsines se trouvent sur le
chromosome 7 pour l’opsine B, et sur le chromosome X pour les opsines R et V.
Chez les Primates, les grands singes possèdent les trois types d’opsine alors que la plupart des autres
Primates n’en possèdent que deux (B et R).
L’analyse des séquences géniques montre la grande ressemblance entre les opsines R et V : il y a eu
duplication du gène R puis évolution des deux gènes qui ont chacune accumulé des mutations et ont
donc permis la formation de deux protéines aux propriétés différentes.
Les séquences nucléotidiques de la rhodopsine et des trois opsines montrent également des
similitudes, moins marquées qu’entre les opsines R et V : les opsines et la rhodopsine forment une
famille multigénique, un gène ancestral ayant été dupliqué, soit sur le même chromosome, soit sur un
chromosome différent, chaque copie ayant ensuite accumulé des mutations ponctuelles indépendamment
des autres et donc acquis des propriétés différentes.
La comparaison du nombre d’opsines, et des séquences géniques, est l’un des moyens de reconstituer
l’évolution des Mammifères et en particulier des Primates, et confirme la forte parenté entre l’espèce
humaine et les grands singes.
IV. De la rétine au cerveau
Répondre aux questions p 313
1. Zone occipitale
L’image, obtenue par tomographie par émission de positon (TEP) lorsqu’un individu lit des mots,
montre que l’arrière du cerveau ainsi qu’une zone située sur la partie arrière droite sont actives. Ce sont
les seules zones actives, et elles diffèrent de celles actives lors d’une stimulation auditive.
La zone occipitale (arrière du cerveau) qui est activée par des stimulations visuelles est une cartographie
du champ visuel : chaque secteur de cortex correspond à une partie du champ visuel. Les zones du
champ visuel correspondant à la fovéa sont représentées dans le cerveau par une zone très étendue.
Schéma p 317
2. Trajet du message nerveux.
Lorsqu’une cellule photoréceptrice est excitée par la lumière, la molécule de rétinal constituant une
partie de l’opsine ou de la rhodopsine change de configuration : de configuration cis en l’absence de
lumière, elle passe en configuration trans. Ce changement de configuration modifie l’activité électrique

4
de la cellule photoréceptrice, provoquant la naissance d’un message nerveux transmis aux cellules
bipolaires puis aux cellules ganglionnaires. Le message est ensuite acheminé vers les aires visuelles du
cortex cérébral.
Au retour à l’obscurité, une enzyme assure la transformation inverse de la forme trans à la forme cis du
rétinal.
Les nerfs optiques se croisent au niveau du chiasma des nerfs optiques. Des sections opérées à différents
niveaux montrent que :
- Avant le chiasma, les nerfs optiques transmettent les messages visuels en provenance de l’œil dont ils
sont issus.
- Au niveau du chiasma, la plupart des fibres nerveuses optiques se croisent : les messages visuels en
provenance de l’œil droit sont transmis à l’hémisphère gauche du cerveau et vice versa. Cependant les
fibres en provenance de la partie de la rétine côté interne ne se croisent pas.
1
/
4
100%