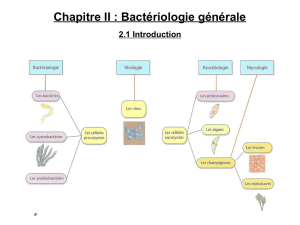
Bactéries à multiplication intracellulaire

Immunologie – L’immunité antibactérienne – page 1/5
Marion et Guigui
31/03/09
IMMUNOLOGIE
15H à 16H
M&M's
Sev Boullier
L’immunité anti bactérienne (suite).
Immunopathologie induite par les bactéries
On va s’intéresser aux phénomènes immunopathologiques induits par les bactéries et voir comment
malgré les mécanismes de protection, les bactéries peuvent échapper à la réponse immunitaire.
HSI : jusqu’à présent aucun phénomène de HSI n’a été décrit et dans aucune espèce animale, à la
suite d’une infection bactérienne.
HSII : elle est caractérisée par la destruction de cellules à la suite de la reconnaissance d’un
antigène à sa surface.
On a déjà vu précédemment que le LPS (très immunogène) se trouve à la surface des bactéries G-.
Celui-ci entraîne une activation massive des macrophages, responsable d’un choc septique et peut
également s’effeuiller et se retrouver libre dans la circulation sanguine. Libre, il a une très forte
affinité pour les globules rouges, sur lesquels il s’adsorbe. Or ce LPS est associé à une réponse de
type humorale ; les AC anti-LPS produits vont se fixés sur les LPS adsorbés à la surface du globule
rouge entraînant leur destruction. On a une HSII avec une lyse massive des globules rouges,
provoquant anémie et choc inflammatoire. Ceci fera l’objet d’un diagnostic différentiel avec
certaines anémies hémolytiques très brutales.
Rq: Choc septique: choc de
l’organisme dû à une infection,
activation massive de la réaction
inflammatoire. On met en place un
traitement pour limiter
l’inflammation à base de corticoïde
auquel on ajoute une couverture anti
bactérienne.
HSIII : Elle est associée au dépôt des immun-complexes sur les membranes biologiques, elle
résulte généralement d'une combinaison entre une réponse humorale et une infection chronique.
D’une manière générale, toute infection bactérienne avec une multiplication extracellulaire et durant
plus de 3semaines peut être associée à une HSIII.
Ex : phase chronique de la gourme du cheval (Streptococcus equi). C’est l’exemple le plus
classique. Il s’agit d’une angine à streptococcus equi. L’efficacité des antibiotiques est limité sur
cette infection bactérienne. Elle peut entraîner des pétéchies, une insuffisance rénale chronique…
L’animal peut rester porteur de la bactérie pendant plusieurs mois.
HSIV : lorsque l’on a des bactéries à multiplication intracellulaire dans les macrophages, et
responsables de la formation de granulomes (cf tuberculose). Cette hypersensibilité est associée à
une infection chronique avec une forte réponse cellulaire.

Immunologie – L’immunité antibactérienne – page 2/5
Mécanismes d’échappement à la réponse immunitaire
Certaines bactéries sont résistantes malgré la réponse extrêmement performante. Elles ont
développé des techniques d’échappement à cette réponse.
Les bactéries à multiplication intracellulaire ne développent pas les mêmes techniques que les
bactéries à multiplication extracellulaire.
Résistance à l’attaque par des facteurs chimiques
Notamment pour les bactéries à multiplication extracellulaire car elles sont plus sujettes à
l’agression par les facteurs chimiques.
Staphylococcus aureus est résistant au lysozyme et aux défensines du fait de la constitution de sa
paroi et la présence d’inhibiteurs spécifiques qu’il produit. Ces molécules ne peuvent donc pas faire
de trou dans la paroi des Staphylococcus.
Dans le mucus on a une concentration importante de ces facteurs chimiques, or certaines bactéries
ont acquis la capacité à dégrader le mucus et ainsi arriver plus facilement au niveau des muqueuses
et s’accrocher sur les cellules épithéliales. (ex : Streptococcus pneumoniae).
Inhibition de l’activité du complément
Les bactéries à multiplication extracellulaire, pour ne pas être détruites rapidement, doivent
échapper au complément (l’activation du complément entraîne destruction cellulaire ou
recrutement des granulocytes neutrophiles).
Ainsi, certaines bactéries G- voient une modification de leur paroi, et ne seront donc pas reconnues
par la voie alterne du complément. En revanche, elles seront toujours « visibles » par la voie
classique. Mais celle-ci est plus longue à mettre en place. Il y a donc un retard de la réponse.
Certaines bactéries peuvent recruter sur leur paroi des inhibiteurs du complément. En effet, les
cellules eucaryotes possèdent à leur surface ces inhibiteurs du complément (afin de ne pas être
détruites lors de son activation (activité très peu spécifique)).
Ex: Les pneumocoque recrutent le facteur H.
On note qu'il n'y a pas de destruction directe et pas non plus de production de facteurs
chimiotactiques et inflammatoires
Résistance à l’opsonisation
L’opsonisation est la meilleure arme pour éliminer les bactéries extracellulaires, qui seront
phagocytées par les macrophages ou les granulocytes. Certaines bactéries G+ possèdent une capsule
qui limite leur destruction, mais aussi la présentation antigénique.
Ainsi, les cellules dendritiques ont plus de mal à phagocyter ces bactéries et à présenter les
antigènes. La réponse spécifique est donc moins bonne (car moins bonne présentation).
Les bactéries pathogènes extracellulaires ont déjà éliminé une bonne partie des mécanismes non
spécifiques.

Immunologie – L’immunité antibactérienne – page 3/5
Résistance à la lyse par les cellules phagocytaires
Ce mécanisme met en jeu les macrophages ou les granulocytes.
Il y a toujours une bactérie pour y échapper. Le mécanisme de résistance à l’attaque des
granulocytes peut se faire aussi bien en milieu intracellulaire qu’en milieu extracellulaire.
Résistance dirigée contre les granulocytes
Bactéries à multiplication extracellulaire.
Les bactérie sont en général détruites par les granulocytes. En cas de résistance, la bactérie persiste
dans l’organisme. Cette résistance peut être due à:
-Une inhibition de la phagocytose (E coli);
-La production de toxines lytiques (Streptolysine, leucocidines), favorisant la production de
pus;
-L'inhibition de la mobilité des granulocytes neutrophiles. Les staphylocoques par exemple
peuvent synthétiser des facteurs empêchant la diapédèse des GNN (granulocytes neutrophiles), ces
derniers restent donc coincés dans les vaisseaux. Les bactéries vont donc subir la phagocytose mais
ne vont pas être détruites.
-La résistance aux dérivés oxygéné: Les GNN produisent des radicaux oxygènes, et
certaines bactéries ont mis en place des systèmes de lutte contre ces radicaux: les catalases, qui
empêche leur production (si vous vous rappelez de nos TP de l’année dernière en immunologie, il
reviendra peut être à votre esprit que nous avions testé la caractéristique catalase + (ou -) avec nos
petites galeries API… ça vous revient… ben maintenant grâce à Séverine vous êtes un peu plus
renseignez sur le sujet).
-L'attraction et activation locale des GNN et résistance à la phagocytose:
Une bactérie résistante détourne le système immunitaire à son profit. Un petit exemple,
Helicobacter pylori, (responsable de 50% des ulcères chez l’homme). La bactérie se développe dans
l’estomac ,en résistant au pH acide de celui-ci. La bactérie produit elle même des facteurs qui
recrutent les GNN sur le site infectieux et leur activation. Ce phénomène favorise le développement
d’ulcère. De plus cette bactérie synthétise des facteurs détruisant les GNN, ce qui lui fournit ses
propres aliments (déchets des GNN qu’elle a elle même recruté). Ces bactéries ne peuvent donc pas
être éliminées sans traitement.
Bactéries à multiplication intracellulaire.
Il existe des bactéries qui se multiplient dans les GNN, comme certaines anaplasma (ehrlichiose
chez l’homme)
Elles provoquent un blocage de l’explosion respiratoire, en bloquant toute la cascade de la
production des radicaux oxygénés ; cela bloque le fonctionnement des granules des GNN. Il y a
alors une baisse générale des activités bactéricides des GNN. Les GNN deviennent des cellules
désarmées.
Résistances dirigées contre les macrophages:
Bactéries à multiplication intracellulaire
Ce type de résistance a été mis en place par des bactéries tel que Brucella et certaines
Mycobactéries qui se multiplient dans les macrophages. Ce type de bactérie désactive la fonction
bactéricide des macrophages. Les systèmes de résistance développés sont :
-Résistance aux enzymes lysosomiales (corynebactérium, legionella). Les macrophages
possèdent des vacuoles lysosomiales à enzymes a priori extrêmement puissantes.

Immunologie – L’immunité antibactérienne – page 4/5
-Blocage de la fusion du phagosome avec le lysosome (MTB, brucella). La multiplication
des bactéries dans le phagosome empêche la fusion de ce dernier avec les lysosomes par une
modification des filaments dans le cytoplasme.
-Echappement du phagosome avant la fusion et multiplication dans le cytoplasme. Les
bactéries utilisent la vacuole de phagocytose pour entrer dans le macrophage, puis sortent de la
vacuole avant la fusion avec les lysosomes. Les bactéries se multiplient alors dans le cytoplasme du
macrophage (c’est le cas de Listeria). Ces bactéries peuvent être responsable de dysentries sévères,
elles induisent la mort par apoptose du macrophage (c’est le cas de Shigella).
Inhibition de la réponse spécifique
Il existe différent mécanisme d’inhibition de la réponse immunitaire spécifique :
-Inhibition de l’apoptose (shigella). La bactérie entraîne un blocage de l’apoptose de la
cellule infectée, les cellue cytolytique n’ont aucune efficacité.
-Inhibition de l’activité des IgA:
On distingue: Protéases IgA spécifiques: On a découvert des protéases de bactéries spécifiques des
IgA (qui avaient pourtant la particularité de résister aux protéases). Ces bactéries peuvent donc se
multiplier au niveau des muqueuses (ex : Neisseiria, pneumocoques).
Leurres FcRα (Staphylococcus), ce sont des récepteurs, leurres immunologiques.
-Modification de l’activité des cytokines: notamment TNF : reconnaissance TNF-R
(protéine A staph). Cette modification des cytokines est très forte lors d’une infection à
staphylocoques. Les protéines A empêchent la fixation normal du TNF sur son récepteur.
-Blocage des FcR: (protéine A staph). D’une façon générale, les protéines A des
staphylocoques bloquent tout les Fc récepteurs.
-Inhibition de l’expression des CMH-II (helicobacter pylori). S’il y a blocage de
l’expression des CMH-II, il ne peut y avoir mise en place d’une réponse immunitaire induite
-Production de super Antigènes (Staph et staph. doré). Ces super antigènes sont à l’origine
d’une activation massive, non spécifique des LT (lymphocyte T). Cette activation est incontrôlée,
les lymphocytes meurent de cette stimulation anormale. Ceci entraîne une immunodépression.
Modifications du profil de la réponse spécifique
Mycobacterium leprae: forme lépromateuse ou tuberculoïde de la lèpre
Cette bactérie induit l’activation des cellules présentatrices d’antigène vers un profil Th2, il n’y a
donc pas des interférons γ. Or, elle se multiplie dans les macrophages. La lutte contre celle-ci
nécessite donc des interférons γ. On parle alors de deux lèpres.
-la forme lépromateuse : il y a une dissémination massive du bacille dans l’organisme, pas
de granulome formé et aucun contrôle de la multiplication bactérienne. C'est une infection
rapidement mortelle.
-la forme tuberculoïde : c’est la forme classique (de résistance) de la lèpre. La formation de
granulomes bloque la diffusion du bacille et la circulation sanguine et nerveuse, ce qui explique les
nécroses des extrémités. Les traitements sont de nos jours efficaces contre cette forme.
Variation antigénique
Ce processus est beaucoup moins important que pour les virus. Ce système d’échappement
concerne différentes bactéries.
Spirochètes, Campylobacter fetus, streptococcus pneumoniae
Récemment on s’est aperçu que Strep. pneumoniae peut varier ces facteurs de virulence au cours de
la réponse immunitaire.

Immunologie – L’immunité antibactérienne – page 5/5
Diagnostic des infections bactériennes
Le diagnostic peut être direct ou indirect.
Direct : Recherche de la présence d’antigène par coloration, culture ou PCR.
Il faut toujours garder en mémoire qu’il peut y avoir des faux négatifs et des faux positifs. Les faux
positifs en diagnostic direct représentent les cas ou la bactérie présente n’est pas responsable de
signes cliniques (ex : présence de salmonella dans le tube digestif d’un cheval sans aucun signe
clinique).
Rq: Présence de bactérie ne signifie pas forcément signe clinique.
Attention présence bactérie≠virulence
Indirect : Détection de marqueurs spécifiques de la réponse immunitaire, indiquant la présence
d’antigène.
In vivo: mise en évidence de la réponse à médiation cellulaire.
In vitro: sérodiagnostic principalement.
Il faut savoir identifier les faux positifs et les faux négatifs!
Il ne faut pas oublier la fonction mémoire du système immunitaire : détecter un marqueur ne veut
pas dire que l’antigène est toujours présent.
Il existe des faux positifs pour les infections non chroniques, et des faux négatifs dû à la
séroconversion, à la sensibilité de la technique ou à une immunodépression passagère.
1
/
5
100%