PDF(10,27Mo) - Collection mémoires et thèses électroniques


Étude du rôle de la voie ERK/MAPK dans le
développement embryonnaire chez la souris
Thèse
Rifdat Aoidi
Sous la direction de
Jean Charron, directeur de recherche

iii
Résumé
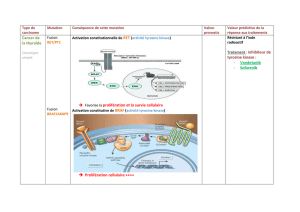
Les mammifères possèdent deux MAP kinases kinases (MEK1 et MEK2), impliquées dans
l’activation de la voie ERK/MAPK essentielle pour la différenciation, la prolifération et la survie
cellulaire. Le premier objectif de cette thèse était de déterminer si les fonctions des kinases MEK1 et
MEK2 sont redondantes durant le développement embryonnaire. Les souris Mek1-/- meurent à mi-
gestation d’une malformation du placenta. Les souris Mek2-/- ne présentent aucun phénotype majeur,
suggérant que ces deux protéines ont des rôles différents. Cependant, la plupart des mutants Mek1+/-
Mek2+/- meurent pendant la gestation d’un sous-développement du placenta, indiquant que Mek1 et
Mek2 ont chacun un rôle dans le développement des tissus extraembryonnaires. À ce jour aucune
évidence claire ne permet de statuer sur la redondance fonctionnelle de MEK1 et MEK2. Afin de
vérifier la spécificité fonctionnelle de Mek1 et Mek2, nous avons généré au laboratoire un allèle
« knockin », exprimant l’ADNc de Mek2 sous contrôle du locus Mek1 (Mek12). L’analyse de ces
souris a révélé la redondance fonctionnelle entre MEK1 et MEK2. L’analyse de combinaisons
alléliques de Mek a démontré qu’une expression minimale de protéines MEK est cruciale pour le
développement embryonnaire et la survie.
Le second objectif de cette thèse était de caractériser les mutants Mp1. Les protéines
d’échafaudage permettent de moduler l’activité de la voie ERK/MAPK et facilitent la transmission
rapide du signal. Parmi les protéines d’échafaudage connues, seule MP1 (Mek Partner 1) a été
identifiée comme étant un partenaire spécifique de MEK1 et ERK1. Cette spécificité suggère que
MP1 pourrait contribuer à la différence d’activation de MEK1 et MEK2 en spécifiant le signal qui
passe par Mek1. Afin d’étudier le rôle de Mp1 au cours du développement chez la souris, nous avons
généré des souris Mp1-/-. L’analyse de ces mutants indique que le gène Mp1 est essentiel pour la
survie et que sa fonction est nécessaire suite à la post-implantation.
La dérégulation de la voie ERK/MAPK dans le développement chez l’homme a aussi des
conséquences phénotypiques. Au cours des dernières années, une classe de syndromes a été
caractérisée : Les « Rasophaties ». Ces syndromes partagent des caractéristiques communes qui
sont, une mutation dans des gènes de la voie ERK/MAPK, une dysmorphologie cranio-faciale, des
malformations cardiaques et cutanées ainsi qu’un retard mental. Parmi les mutations de la voie
ERK/MAPK qui ont été identifiées, une mutation ponctuelle dans le gène Mek1 (Mek1Y130C) cause le
syndrome Cardio-Facio-Cutané (CFC). Le dernier objectif de cette thèse était de générer un modèle
animal pour le CFC portant la mutation Mek1Y130C. Les souris portant l’allèle Mek1Y130C présentent les

iv
phénotypes associés au CFC (i.e sténose pulmonaire, dysmorphologie cranio-faciale et défauts
neurologiques).

v
Abstract
Mammals possess two MAP kinase kinase (MEK1 and MEK2), involved in ERK/MAPK pathway. This
pathway is essential for proliferation, differentiation and cell survival. The first objective of my thesis was to
determinate if MEK1 and MEK2 kinases are redundant during embryonic development. Mek1-/- mice die at
embryonic day E10.5 due to placental defects, whereas Mek2-/- mice survive with a normal lifespan
suggesting that MEK1 possesses functions not shared by MEK2. However, most Mek1+/-Mek2+/-
embryos also die from placental defects, indicating that both Mek genes contribute to placental
development. To date, no clear evidence on MEK1 and MEK2 redundancy has been provided. To assess
the functional specificity of the Mek1 and Mek2 genes, we produced a Mek1-knockin allele in which
the Mek2 coding sequences were placed under the control of Mek1 regulatory sequences. Analyzing
these mice allowed us to demonstrate that MEK1 and MEK2 can substitute for each other and that a
minimal amount of MEK is critical for placenta development and embryo survival.
The second objective of my thesis was to characterize Mp1 mutants. Scaffold proteins
modulate MAPK pathway by providing spatial and temporal specificity. Among known ERK/MAPK
scaffold proteins, only MP1 (Mek Partner 1) is specific to MEK1 and ERK1, raising the question of the
specificity of MP1 in the regulation of ERK/MAPK pathway via MEK1. In order to investigate Mp1
function in vivo, we generated Mp1 knock-out mice. Analyzing these mice enable us to suggest that
Mp1 is required for embryonic development and is essential during post-implantation.
Deregulation of Ras/MAPK pathway also causes developmental phenotypes in human.
During the last decade, a new class of syndromes, which share common phenotypes such as
mutations in Ras/MAPK pathway, cranio-facial dysmorphology, cardiac and cutaneous malformations
and neurological delay has been described and named Rasophaties. Among the DNA mutations
found in rasopathies, the Mek1 mutation, Mek1Y130C, causes cardio-facio-cutaneous syndrome (CFC).
The last objective of my thesis was to generate a mouse model of CFC, with the Mek1Y130C mutation.
I found that mice carrying the Mek1Y130C mutation partially recapitulate CFC syndrome (i.e pulmonary
stenosis, crani-facial dysmophia and neurological defects).
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
1
/
276
100%