I. Le cristallin, une lentille vivante - Les SVT à Stella

Première S - Cours 3
DE L’ŒIL AU CERVEAU – QUELQUES ASPECTS DE LA VISION
I. Le cristallin, une lentille vivante :
1°) Les parties de l’œil en relation avec la
formation d’une image :
Voir la dissection de l’œil de thon.
Schéma représentant les
différentes parties de l’œil
Il est indispensable de revoir le cours d’optique
fait en physique
2°) Organisation structurale du cristallin :
Voir la suite du TP sur l’œil de thon
Cristallin observé au microscope x400
Le cristallin est enfermé dans une capsule mince et transparente et maintenu par les ligaments
suspenseurs. Il est organisé en couches de cellules fibreuses.
Ces cellules n'ont pas de noyau, et très peu d'organites (contrairement aux autres cellules
animales). Elles renferment des protéines : les cristallines qui sont transparentes et ont une
activité enzymatique permettant de produire de l'énergie à partir des sucres.
L'absence d'organites explique leur transparence. Les substances passent par simple diffusion
d’une cellule à l’autre.
Le cristallin n'est ni vascularisé, ni innervé.
Les cellules du cristallin sont pour la plupart créées pendant la vie embryonnaire et
presqu'aucune ne se forme après l'âge de 20 ans (elles n’ont pas de noyau et ne peuvent
donc pas se diviser)
Muscle ciliaire
Ligament
Cristallin
Cornée
Humeur
aqueuse
Iris
Epithélium
Capsule
antérieure
Sutures
Cellules en ruban
du cortex et du
noyau
Capsule
postérieure
Face
antérieure
Face
postérieure

3°) Propriété d’accommodation du cristallin :
Le cristallin est souple et déformable. Grace aux ligaments et muscles qui le maintiennent en
place, il peut changer de courbure et ainsi réaliser l'accommodation, c'est à dire la vision nette
des objets plus ou moins proche.
4°) Défauts de vision en relation avec l’atération du cristallin :
Si un défaut de courbure présent dès la naissance est à l’origine de problème de vision comme la
myopie et l’astigmatisme (voir cours d’optique), un vieillissement du cristallin provoque d’autres
manifestations : La structure du cristallin n'étant pas renouvelée. Au fil du temps, cet organe se
détériore et peut provoquer des défauts de vision.
- La presbytie : Les cellules du cristallin sont moins souples avec l'âge, elles se déforment moins
et donc la capacité d'accommodation du cristallin diminue (l’image nette se forme en arrière
de la rétine) Il faut donc porter des lunettes pour voir des images nettes de près.
- La cataracte : C'est une opacification du cristallin du à la précipitation irréversible des
protéines du cytoplasme.
Il est possible de se faire opérer de la cataracte (on remplace le cristallin défaillant par un
implant artificiel) Cependant c'est la première cause de cécité dans le monde.
Les rayons lumineux traversent donc les milieux transparents de l’œil et convergent pour former
une image nette au niveau de la rétine.
De quoi est formée cette membrane sensible à la lumière ?
II. La rétine, une membrane photosensible :
1°) Organisation structurale de la rétine :
Voir dessin d’observation ;
La rétine est formée de trois couches de
cellules nerveuses.
En partant de l’humeur vitrée, on a d’abord les
neurones ganglionnaires, puis les neurones
bipolaires, et enfin les cellules photo-
réceptrices. Les cellules photosensibles sont
en contact avec la choroïde.
Il existe deux types de cellules photo
réceptrices différentes anatomiquement.
Les bâtonnets et les cônes.

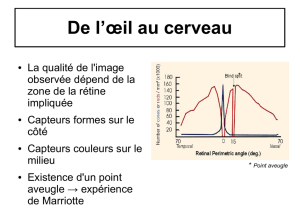
2°) Répartition, rôle et composition des photorécepteurs
Les bâtonnets sont des photorécepteurs extrêmement
sensibles à la lumière. Ils sont très nombreux au niveau de la
rétine périphérique, ils permettent de voir dans des conditions
d'éclairage faible et assurent ainsi la vision crépusculaire en
nuances de gris.
Les cônes sont concentrés dans l'axe de l'œil (fovéa = environ
1,5 mm de diamètre)
Il en existe trois types différents sensibles à la lumière
rouge, verte, et bleue. Les informations recueillies par les
trois types de cônes permettent de percevoir les couleurs par
synthèse additive. Les cônes assurent donc la vision diurne
(vision de jour) en couleurs.
La mesure du champ visuel permet de comprendre la répartition des cônes. C’est la zone totale
dans laquelle la perception visuelle est possible pour une couleur donnée quand la personne
regarde devant elle.
Les photorécepteurs sont capables de convertir la stimulation lumineuse en un message nerveux
de nature électrique. Ce message sera traité par les cellules bipolaires et ganglionnaires puis
transmis au cerveau.
Au niveau de la rétine périphérique, l'information est "compressée" en raison de la convergence des cellules
(beaucoup de photorécepteurs connectés à un petit nombre de cellules ganglionnaires). Dans la zone centrale,
chaque photorécepteur est connecté à une cellule ganglionnaire : l'acuité est alors maximale. Dans la zone
centrale, chaque photorécepteur est connecté à une cellule ganglionnaire : l'acuité est alors maximale.
Les photorécepteurs contiennent dans leur segment externe, des structures riches en pigment
rétinien. Les pigments visuels contenus dans les cônes sont constitués d'une protéine, l'opsine.
Il y a 3 sortes de cônes contenant chacune une opsine
sensible à une longueur d’onde donnée :
Les opsines L sensibles à la couleur rouge. Les opsines
M sensibles à la couleur verte. Les opsines S sensibles
à la couleur bleue.
Le pigment visuel contenu dans les bâtonnets est
constitué d'une protéine, la rhodopsine.
Si une de ces molécules n’est pas sensible à sa couleur
(protéines dysfonctionnelles ou non-sens) l’individu souffre de
défauts de vision des couleurs. C’est le daltonisme. C’est une
déficience génétique.
Dans le cas le plus courant, les individus ne peuvent pas
synthétiser une opsine L fonctionnelle et sont donc
dichromates. Les rouges leur paraissent sombres et ils
distinguent mal le vert, le jaune et l’orange.
La déficience des photorécepteurs peut avoir d’autres origines :
- liée à l'âge, comme dans la dégénérescence maculaire (DMLA) due à une défaillance des cônes et qui
entraîne une perte de la vision centrale ;
- alimentaire, comme dans l'héméralopie, qui est une défaillance en vitamine A qui entraîne une perte de la
vision nocturne.

3°) Relations de parenté entre les molécules des pigments rétiniens :
La synthèse des pigments rétiniens est
gouvernée par des gènes portés par les
chromosomes n°7 et X.
L’observation des séquences de ces
protéines (ou des gènes correspondants)
montre un pourcentage de similitude que le
seul hasard ne peut expliquer.
On parle alors de famille multigénique. Les
gènes qui codent pour ces molécules dérivent
d'un « gène ancestral » commun.
Les molécules et les gènes montrant des
similitudes sont qualifiées d’homologues.
En effet, un gène peut-être accidentellement copié
et se retrouver présent dans le génome en deux
exemplaires : c'est la duplication. Certaines copies
peuvent également subir un déplacement sur un
autre chromosome : c'est une transposition.
Par la suite, des mutations ponctuelles se produisent et
rendent différents ces duplicata initialement identiques.
Plus la duplication d'un gène est ancienne et plus les deux
gènes qui en résultent sont différents.
Ces gènes peuvent permettre la production de protéines
remplissant des fonctions différentes.
4°) Utilisation les opsines pour établir des relations de parenté entre les primates:
L’étude des séquences des opsines chez différents primates comparés à l’Homme indique que :
- Certaines opsines sont absentes chez certaines espèces.
- La même molécule (opsine bleue) montre des différences entre les espèces mais avec des
degrés de similitudes différents.
Ainsi, tous les primates sont
apparentés car leur gène qui code
pour l’opsine sensible au bleu
présente de grandes similitudes. Le
degré de similitude et la présence de
toutes les opsines permettent
d’affiner cette parenté.
On peut donc conclure qu’il s’est
produit, au sein du groupe des
primates, un phénomène
d’enrichissement du génome, à la
base de la vision trichromatique qui caractérise l’Homme et les primates proches.
L’étude comparée des pigments rétiniens montre donc que la vision est bien un produit de
l’évolution des espèces.

III. De la rétine au cerveau :
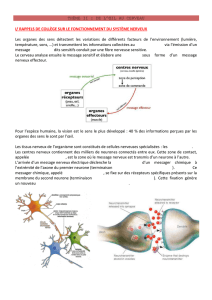
1°) Le message nerveux visuel:
Les cellules photosensibles contiennent des
pigments (rhodopsine et opsines) capables de
transformer un signal lumineux en signal électrique
(=il se forme dans la cellule une dépolarisation)
Ce signal électrique provoque la naissance d’un
message nerveux dans les cellules bipolaires qui
sera ensuite transmis aux cellules ganglionnaires
dont le prolongement permet au message de sortir
de l’œil par le nerf optique.
Le message nerveux est constitué d’un ensemble de
potentiels d’action (=variations brusques de la
tension dont l’amplitude est toujours la même)
C’est la fréquence des PA qui constitue un
message (voir doc livre page 313)
2°) Régions du cerveau impliquées dans la vision :
Le cerveau comporte deux hémisphères cérébraux. On distingue 4 grands lobes à la surface de chaque
hémisphère, les lobes frontal, pariétal, temporal et occipital. Chaque
hémisphère comporte une mince couche de matière grise externe, appelée
cortex cérébral. Du fait des nombreux replis ou circonvolutions cérébrales,
le cortex représente 40% de la masse de l’encéphale (cerveau, cervelet et
bulbe rachidien).
Il est constitué par l’assemblage de plusieurs dizaines de milliers de
cellules nerveuses ou neurones.
L’Imagerie par Résonance Magnétique nucléaire (IRM) est une technique
inoffensive qui permet d’observer des coupes virtuelles de l’organisme
avec une résolution qui peut atteindre quelques micromètres.
L’IRMf (=fonctionnelle) permet
de visualiser les zones dont
l’activité est importante.
On observe que l’arrivée du message nerveux visuel
provoque l’activation d’une partie du lobe occipital
appelée cortex visuel primaire.
D’autres zones corticales proches entrent ensuite en activité ce
qui laisse penser que la fonction visuelle mobilise des zones
corticales associées comme le lobe pariétal
3°) Voies visuelles entre l’œil et le cerveau :
L’exploration du champ visuel permet l’étude des voies visuelles qui vont de la rétine au cortex visuel
occipital. Il est en effet possible de mettre en relation des déficits du champ visuel avec l’étude des
conséquences de lésions des voies visuelles observées chez certains individus.
Les voies visuelles sont constituées des nerfs optiques qui se rejoignent au niveau d’un chiasma.
La continuation des nerfs optiques après le chiasma s'appelle le tractus optique. Dans chaque
hémisphère, il transfère l'information visuelle jusqu’au cortex visuel en passant par différents
différentes zones relais appelées noyaux.
 6
7
8
6
7
8
1
/
8
100%